Tagasikliinik Kliinilise neurofüsioloogia tugi. El Paso, TX. Kiropraktikko, Dr. Alexander Jimenez arutab kliiniline neurofüsioloogia. Dr Jimenez uurib perifeersete närvikiudude, seljaaju, ajutüve ja aju kliinilist tähtsust ja funktsionaalset toimet vistseraalsete ja lihas-skeleti haiguste kontekstis. Patsiendid saavad arenenud teadmisi anatoomia, geneetika, biokeemia ja valu füsioloogia kohta seoses erinevate kliiniliste sündroomidega. Kombineeritakse natsitseptsiooni ja valu seotud toitumisharjumusi. Ja rõhutatakse selle info rakendamist raviprogrammidesse.
Meie meeskond tunneb suurt uhkust selle üle, et toovad oma peredele ja vigastatud patsientidele ainult tõestatud raviprotokolle. Õpetades terviklikku terviklikku heaolu kui elustiili, muudame me mitte ainult oma patsientide, vaid ka nende perede elusid. Teeme seda selleks, et jõuaksime võimalikult paljude El Pasolasteni, kes meid vajavad, olenemata taskukohasuse probleemidest. Kõigile teie küsimustele vastuste saamiseks helistage dr Jimenezile numbril 915-850-0900.
"Kliinilised otsustusreeglid, seljavalu klassifikatsioon ja ravitulemuste prognoosimine: arutelu hiljutiste aruannete üle rehabilitatsioonikirjanduses"
Abstraktne
Kliiniliste otsuste reeglid on biomeditsiinilises kirjanduses üha levinumad ja need on üks strateegia kliiniliste otsuste tegemise tõhustamiseks, et parandada tervishoiuteenuste osutamise tõhusust ja tulemuslikkust. Rehabilitatsiooniuuringute kontekstis on kliiniliste otsuste reeglid olnud peamiselt suunatud patsientide klassifitseerimisele, ennustades nende ravivastust konkreetsetele ravimeetoditele. Traditsiooniliselt on kliiniliste otsuste reeglite väljatöötamise soovitustes ette nähtud mitmeastmeline protsess (tuletamine, valideerimine, mõjuanalüüs), kasutades määratletud metoodikat. Diagnoosil põhineva kliinilise otsuse reegli väljatöötamiseks tehtud teadusuuringud on sellest kokkuleppest kõrvale kaldunud. Selle uurimissuuna hiljutistes väljaannetes on kasutatud muudetud terminoloogia diagnoosipõhist kliiniliste otsuste juhendit. Kliiniliste otsuste reeglitega seotud terminoloogia ja metoodika muudatused võivad arstidel raskendada otsustusreegliga seotud tõendite taseme äratundmist ja mõista, kuidas neid tõendeid tuleks patsiendihoolduse teavitamiseks rakendada. Anname lühikese ülevaate kliiniliste otsuste reeglite väljatöötamisest rehabilitatsioonialase kirjanduse ja kahe konkreetse dokumendi kontekstis, mis avaldati hiljuti ajakirjas Chiropractic and Manual Therapies.
Kliinilise prognoosimise reeglid
Tervishoid on läbi teinud olulise paradigma muutuse tõenduspõhise praktika suunas. Arvatakse, et lähenemisviis parandab kliiniliste otsuste tegemist, integreerides parimad saadaolevad tõendid kliiniliste teadmiste ja patsientide eelistustega.
Lõppkokkuvõttes on tõenduspõhise praktika eesmärk parandada tervishoiuteenuste osutamist. Teaduslike tõendite praktikasse tõlkimine on aga osutunud keeruliseks ettevõtmiseks.
Kliiniliste otsuste reeglid (CDR-id), mida tuntakse ka kliiniliste prognooside reeglitena, on rehabilitatsioonialases kirjanduses üha tavalisemad.
Need on vahendid, mis on loodud kliiniliste otsuste tegemisel teavitamiseks, tuvastades diagnostiliste testide tulemuste, prognoosi või ravivastuse potentsiaalsed ennustajad.
Taastusravi kirjanduses kasutatakse CDR-e kõige sagedamini patsiendi ravivastuse ennustamiseks. Neid on pakutud välja selgitada kliiniliselt olulised patsientide alarühmad, kellel esinevad muidu heterogeensed häired, nagu näiteks mittespetsiifiline kael või madal. seljavalu, mis on vaatenurk, millele kavatseme keskenduda.
Kliinilise prognoosimise reeglid
Võimalus klassifitseerida või alarühmadesse rühmitada heterogeensete häiretega, näiteks seljavaluga patsiente, on esile tõstetud uurimistöö prioriteedina ja sellest tulenevalt paljude uurimistöö keskmes. Selliste klassifitseerimismeetodite võlu seisneb nende potentsiaalis parandada ravi tõhusust ja tulemuslikkust, sobitades patsiendid optimaalsete ravimeetoditega. Varem on patsientide klassifitseerimine tuginenud kaudsetele lähenemisviisidele, mis põhinesid traditsioonidel või ebasüstemaatilisel vaatlusel. CDR-ide kasutamine klassifitseerimiseks on üks katse tõenditepõhisema lähenemisviisi poole, mis ei sõltu vähem alusetust teooriast.
CDR-id on välja töötatud mitmeastmelise protsessina, mis hõlmab tuletamise, valideerimise ja mõju analüüsi uuringuid, millest igaühel on määratletud eesmärk ja metoodilised kriteeriumid. Nagu kõigi patsientide kohta otsuste tegemiseks kasutatavate tõendite puhul, on asjakohasele uuringumetoodikale tähelepanu pööramine kriitilise tähtsusega, et hinnata rakendamisest saadavat võimalikku kasu.
Kliinilise ennustamise reeglite eelised
See mahutab rohkem tegureid, kui inimese aju suudab arvestada
CDR / CPR mudel annab alati sama tulemuse (matemaatiline võrrand)
See võib olla täpsem kui kliiniline hinnang.
Kliinilise prognoosimise reeglite kliiniline kasutamine
Lõppkokkuvõttes ei seisne CDR-i kasulikkus mitte selle täpsuses, vaid selle võimes parandada kliinilisi tulemusi ja suurendada ravi tõhusust.[15] Isegi kui CDR näitab laiaulatuslikku valideerimist, ei taga see, et see muudab kliiniliste otsuste tegemist või et selle tekitatud muudatused toovad kaasa parema hoolduse.
Selle tekitatud muudatused tagavad parema hoolduse. McGinn jt.[2] tuvastas kolm seletust CDR-i ebaõnnestumisele selles etapis. Esiteks, kui arsti otsus on sama täpne kui CDR-i põhjal tehtud otsus, pole selle kasutamisest kasu. Teiseks võib CDR-i rakendamine hõlmata tülikaid arvutusi või protseduure, mis heidutavad arste CDR-i kasutamast. Kolmandaks ei pruugi CDR-i kasutamine kõigis keskkondades või tingimustes võimalik olla. Lisaks võtaksime arvesse tõsiasja, et eksperimentaalsed uuringud võivad hõlmata patsiente, kes ei esinda täielikult tavahoolduses nähtud patsiente, ja et see võib piirata CDR-i tegelikku väärtust. Seetõttu, et täielikult mõista CDR-i kasulikkust ja selle võimet tervishoiuteenuste osutamist parandada, on vaja pragmaatiliselt hinnata selle teostatavust ja mõju, kui seda kasutatakse reaalset praktikat kajastavas keskkonnas. Seda saab teha erinevate uuringukavadega, nagu randomiseeritud uuringud, klastri-randomiseeritud uuringud või muud lähenemisviisid, näiteks CDR-i mõju uurimine enne ja pärast selle rakendamist.
Klassifitseerimise meetodite levimus lülisamba kahjustusega patsientidel, kasutades McKenzie sündroome, valu sümptomid, manipuleerimise ja stabiliseerumise kliinilise ennustamise reeglid.
Eesmärgid olid (1), et määrata nende nimiväärtushäiretega patsientide osakaalu, kellele McKenzie sündroomide (McK) ja muskaadi diagnoosimise ja teraapia (MDT) hindamismeetodite, manipulatsiooni ja stabiliseerumise kliinilise ennustamise abil võiksid klassifitseerida McKenzie sündroomide reeglid (CPR) ja (2) iga Man CPR-i või Stab CPR-kategooria kohta, määrake McK ja PPC-le klassifikatsiooni levimuse määrad.
CPR-id on keerukad probabilisatsiooni- ja prognostilised mudelid, mille puhul tuvastatud patsiendi omaduste rühm ja kliinilised tunnused ja sümptomid on statistiliselt seotud patsiendi tulemuste mõistliku prognoosimisega.
Teadlased töötasid välja kaks eraldi CPR-i patsientide tuvastamiseks, kes reageerivad manipuleerimisele soodsalt.33,34 Flynn et al. töötas välja algse manipulatsiooni CPR-i, kasutades viit kriteeriumi, st sümptomite puudumine allpool põlve, sümptomite hiljutine ilmnemine (<16 päeva), hirmuvältimise küsimustiku36 madal tööskoor (<19), lülisamba nimmepiirkonna hüpomobiilsus ja puusa sisemine osa. rotatsiooni ROM (>35 vähemalt ühe puusa puhul).33
Flynni CPR-i muutsid seejärel Fritz jt. kahele kriteeriumile, mis hõlmasid sümptomite puudumist allpool põlve ja sümptomite hiljutist ilmnemist (<16 päeva), mis on pragmaatiline alternatiiv, et vähendada arstide koormust esmatasandi arstiabi patsientide tuvastamisel, kes kõige tõenäolisemalt reageerivad tõukejõu manipuleerimisele.34
"Potentia.l kliinilise prognoosimise reeglite lõksud"
Mis on kliinilise prognoosimise reeglid?
Kliiniline ennustusreegel (CPR) on kliiniliste leidude kombinatsioon, mis on statistiliselt näidanud olulist prognoositavust konkreetset ravi saanud patsiendi valitud seisundi või prognoosi määramisel 1,2. CPR-id luuakse mitme muutujaga statistiliste meetodite abil, need on kavandatud kliiniliste muutujate valitud rühmade 3,4, 5 prognoosimisvõime uurimiseks ja on mõeldud selleks, et aidata arstidel teha kiireid otsuseid, mis võivad tavaliselt olla seotud eelarvamustega6. Reeglid on olemuselt algoritmilised ja sisaldavad koondatud teavet, mis tuvastab sihitud seisundi jaoks väikseima arvu statistiliselt diagnostilisi näitajaidXNUMX.
Kliiniliste prognooside reeglid töötatakse tavaliselt välja kolmeastmelise meetodi abil3. Esiteks on CPR-d meid edasiviivalt tuletanud
mitme muutujaga statistiliste meetodite kasutamine kliiniliste muutujate valitud rühmade prognoosimisvõime uurimiseks3. Teine samm hõlmab CPR-i valideerimist randomiseeritud kontrollitud uuringus, et vähendada riski, et tuletusfaasis välja töötatud ennustavad tegurid valiti juhuslikult14. Kolmas samm hõlmab mõjuanalüüsi läbiviimist, et teha kindlaks, kuidas CPR parandab ravi, vähendab kulusid ja määratleb täpselt sihitud eesmärgi14.
Kuigi on vähe arutelu, et hoolikalt konstrueeritud CPR-id võivad kliinilist tava parandada, ei ole minu teada juhtnööre, mis määratleksid infektsioonide CPR-ide metoodilised nõuded kõigile kliinilistele tavadele. Uuringute kavandamise ja aruandluse ranguse parandamiseks on loodud juhised. Järgnevad redaktsioonid kirjeldavad CPR-ide võimalikke metoodilisi lõkse, mis võivad märkimisväärselt nõrgendada algoritmi ülekantavust. Rehabilitatsiooni valdkonnas on enamik CPR-id olnud prescriptiivne; Seega, minu kommentaarid siin peegeldavad kohustuslikke CPR-e.
Metoodilised lõksud
CPR-id on kavandatud määratlema homogeenset omaduste kogumit potentsiaalselt valitud järjestikuste patsientide heterogeensest populatsioonist5,15. Tavaliselt on saadud kohaldatav populatsioon suurema valimi väike alamhulk ja võib moodustada vaid väikese protsendi arsti tegelikust päevasest juhtumite arvust. Suurema valimi seadistus ja asukoht peaksid olema üldistatavad15,16 ning hilisemad kehtivusuuringud nõuavad CPR-i hindamist erinevates patsiendirühmades, erinevates keskkondades ja tüüpilise patsiendirühmaga, mida enamik arste näeb16. Kuna paljud CPR-d on välja töötatud väga erineva rühma põhjal, mis võib, kuid ei pruugi peegeldada tüüpilist patsientide populatsiooni, võib paljude praeguste CPR-algoritmide spektri transporditavus17 olla piiratud.
Kliiniliste prognooside reeglid kasutavad sekkumise tõhususe määramiseks tulemusnäitajaid. Tulemusmeetmetel peab olema üks toimiv määratlus5 ja need peavad nõudma piisavat reageerimisvõimet, et tabada asjakohaseid muutusi tingimuses14; lisaks peaks nendel meetmetel olema hästi üles ehitatud piirhind16,18 ja need peaks koguma pimestatud administraator15. Praegu arutatakse tegelike muutuste mõõtmiseks sobiva ankurdusskoori valikut 19.–20. Enamikul tulemusnäitajatel kasutatakse patsiendi meeldetuletusel põhinevat küsimustikku, nagu globaalne muutuste skoor (GRoC), mis on lühiajalise kasutamise korral asjakohane, kuid kannatab pikaajalistes analüüsides 19–21 kasutamisel tagasikutsumise kõrvalekalde all.
CPR-i võimalikuks puuduseks on suutmatus säilitada algoritmis ennustajatena kasutatud testide ja meetmete kvaliteeti. Seetõttu peaksid perspektiivkatse ja mõõdud olema modelleerimise ajal üksteisest sõltumatud16; igaüks tuleks läbi viia tähendusrikkal ja vastuvõetaval viisil4; arstid või andmehaldurid peaksid olema pimedad patsiendi tulemuste mõõtmise ja seisundi suhtes22.
Allikad
Kliinilise ennustamise reeglite võimalikud lõkse; Manuaalse ja manipuleeriva teraapia ajakiri, 16. köide, number kaks [69]
Jeffrey J Hebert ja Julie M Fritz; Kliiniliste otsuste eeskirjad, spinaalsed valu klassifikatsioonid ja ravi tulemuste ennustamine: arutelu hiljutiste aruannete kohta taastusravi kirjanduses
Depressioon on Ameerika Ühendriikides üks levinumaid vaimse tervise probleeme. Praegused uuringud näitavad, et depressioon tuleneb geneetiliste, bioloogiliste, ökoloogiliste ja psühholoogiliste aspektide kombinatsioonist. Depressioon on kogu maailmas oluline psühhiaatriline häire, mis avaldab ühiskonnale märkimisväärset majanduslikku ja psühholoogilist koormust. Õnneks võib depressiooni, isegi kõige raskemaid juhtumeid, ravida. Mida varem ravi algab, seda tõhusam on see.
Selle tulemusena on siiski vaja püsivaid biomarkereid, mis aitavad parandada diagnoosi, et kiirendada ravimi ja / või ravimi avastamisprotsessi iga haiguspuudega patsiendi jaoks. Need on objektiivsed perifeersed füsioloogilised indikaatorid, mille olemasolu saab kasutada depressiooni tekkimise või olemasolu tõenäosuse ennustamiseks, stratifitseerimiseks vastavalt raskusastmele või sümptomatoloogiale, näidata ennustust ja prognoosi või jälgida ravivastust ravile. Järgmise artikli eesmärk on näidata hiljutisi arusaamu, praeguseid väljakutseid ja tulevikuväljavaateid erinevate avastuste kohta biomarkereid depressiooni ja kuidas need aitavad parandada diagnoosi ja ravi.
Depressiooni biomarkerid: hiljutised teadmised, praegused väljakutsed ja tulevikuväljavaated
Abstraktne
Paljude uuringutega on kaasatud sadu eeldatavaid depressiooni biomarkereid, kuid ei ole veel täielikult välja selgitanud oma rolli depressiivses haiguses või leidnud, milline on ebanormaalne, millistel patsientidel ja kuidas bioloogilist teavet saab kasutada diagnoosi, ravi ja prognoosi parandamiseks. Selline edusammude puudumine on osaliselt tingitud depressiooni olemusest ja heterogeensusest koos teadusliku kirjanduse metodoloogilise heterogeensusega ja potentsiaaliga biomarkerite suur hulgaga, mille ekspressioon muutub sageli erinevate tegurite järgi. Vaatame olemasolevat kirjandust, mis näitab, et põletikuliste, neurotroofsete ja ainevahetusprotsesside, samuti neurotransmitterite ja neuroendokriinsüsteemi komponentidega seotud markerid on väga paljutõotavad kandidaadid. Neid võib mõõta geneetiliste ja epigeneetiliste, transkriptoomsete ja proteoomiliste, metabolomiliste ja neuroimaging hindamiste abil. Uute lähenemisviiside ja süstemaatiliste uurimisprogrammide kasutamine on nüüd vajalik, et teha kindlaks, kas ja milliste biomarkerite abil saab reageerida ravile, stratifitseerida patsiendid spetsiifilistele ravivõimalustele ja arendada uute sekkumiste sihtmärke. Me järeldame, et nende teadustööde edasiarendamise ja laiendamise kaudu on palju lubadusi depressiooni koormuse vähendamiseks.
Märksõnad:meeleoluhäire, suur depressioon, põletik, ravivastus, kihistumine, individuaalne meditsiin
Sissejuhatus
Vaimse tervise ja meeleoluhäirete väljakutsed
Kuigi psühhiaatrias on haiguste tekitatud koormus suurem kui ükskõik milline teine meditsiiniline diagnostikakategooria, on 1 endiselt ilmnenud füüsilise ja vaimse tervise erinevusi paljudes valdkondades, sealhulgas teadusuuringute rahastamine2 ja avaldamine. 3 Vaimse tervisega seotud raskuste seas on puudus klassifitseerimise, diagnoosi ja raviga seotud konsensus, mis tuleneb nende häirete aluseks olevate protsesside ebatäielikust mõistmisest. See ilmneb selgelt meeleoluhäirete kategoorias, mis hõlmab vaimse tervise kõige suuremat koormat. 3 Kõige sagedamini esinev meeleoluhäire, suur depressioonihäire (MDD), on keeruline heterogeenne haigus, mille korral võib kuni 60% patsientidest tekkida mõningane ravivastuse määr, mis pikendab ja halvendab episoode. 4 Meeleoluhäirete ja laiemas vaimse tervise valdkonnas võib ravitulemusi tõenäoliselt parandada, leides tugevaid homogeenseid alatüüpe diagnostikakategooriates (ja nende hulgas), mille abil ravimeetodid võiks olla stratifitseeritud. Selle tunnustamiseks on praegu käimas ülemaailmsed initsiatiivid funktsionaalsete alatüüpide määratlemiseks, näiteks uurimisvaldkondade kriteeriumid. 5 On öeldud, et bioloogilised markerid on vaimsete häirete alltüübi jaoks prioriteetsed kandidaadid. 6
Depressioonravi reageerimise parandamine
Vaatamata ulatuslikule depressiooni ravivõimalustele on ligikaudu kolmandik MDD patsientidest saavutatud remissiooni ka siis, kui võetakse vastu optimaalse antidepressandi ravi vastavalt konsensuse juhistele ja kasutatakse mõõtmispõhist ravi, ning ravivastuse määrad vähenevad iga uue raviga .7 Lisaks on ravile resistentne depressioon (TRD) seotud pikaajalise funktsioonihäirete, suremuse, haigestumuse ja korduvate või krooniliste episoodide suurenemisega. 8,9 Seega võib ravivastuse paranemine igal kliinilisel etapil anda üldise tulemuse laiemale kasule depressioonis. Vaatamata TRD-le tekitatud märkimisväärsele koormusele on selle valdkonna teadusuuringud olnud tagasihoidlikud. Enne TRDde määratlemist ei standardiseerita, hoolimata varasematest katsetest: mõned kriteeriumid 4 nõuavad ainult ühte töötlemisprotsessi, mis ei suuda saavutada 50% sümptomite skoori vähenemist (depressiooni raskusastme valideeritud mõõdetest), samas kui teised eeldavad, et täielik remissioon ei ole saavutatud või reageerimata vastus vähemalt kahele adekvaatselt testitud erinevate klasside antidepressantidele episoodis, mida peetakse TRD.4,10'iks Lisaks parandatakse ravitakistuse määramist ja prognoosimist, lisades ebaõnnestunud ravimeetodite arvu raskusastme ja kroonilatsiooni olulisemad kliinilised tunnused. 9,11 Selle määratluse ebajärjekindlus muudab TRD teadustöö kirjanduse tõlgendamise veelgi keerulisemaks.
Ravile reageerimise parandamiseks on selgelt kasulik tuvastada ennetavaid riskitegureid mittereageerimise korral. On kirjeldatud mõningaid üldiseid TRD ennustajaid, sh täielikku remissiooni puudumist pärast eelnevaid episoode, kaasuvaid ärevust, suitsiidilisust ja depressiooni varajast algust, samuti isiksust (eriti madala ekstraversiooni, madala tasu sõltuvust ja suurt neurotilisust) ja geneetilisi tegureid. 12 Neid avastusi kinnitavad analüüsid, milles sünteesitakse eraldi tõendeid farmakoloogilise 13i ja psühholoogilise 14i depressiooni raviks. Antidepressandid ja kognitiiv-käitumuslikud teraapiad näitavad ligikaudu võrreldavat efektiivsust, 15, kuid nende erinevate toimemehhanismide tõttu võib eeldada, et neil on erinev vastuse ennustaja. Kuigi varajase elu trauma on juba ammu seostatud vaesemate kliiniliste tagajärgedega ja ravivastusega ravivastusega, näitavad 16 varajased näidustused, et lapseea traumaga patsiendid võivad reageerida pigem psühholoogilisele kui farmakoloogilisele ravile. 17 Hoolimata sellest valitseb ebakindlus ja vähe isikupärastamist või ravi stratifitseerimine on jõudnud kliinilisele praktikale. 18
Selles ülevaates keskendutakse tõenditele, mis toetavad biomarkerite kasulikkust kui potentsiaalselt kasulikke kliinilisi vahendeid depressiooni ravivastuse parandamiseks.
Biomarkerid: süsteemid ja allikad
Biomarkerid pakuvad potentsiaalset sihtmärki erinevate interventsioonide reageerimise ennustajate kindlakstegemiseks. 19 Tänapäevased tõendid näitavad, et põletikuliste, neurotransmitterite, neurotroofsete, neuroendokriinsete ja metaboolsete süsteemide aktiivsust peegeldavad markerid võivad ennustada praegusel hetkel depressiooniga inimestel esinevaid vaimseid ja füüsilisi tervise tagajärgi , kuid tulemuste vahel on palju vastuolu. 20 Selles ülevaates keskendume viiele bioloogilisele süsteemile.
Molekulaarsete radade ja nende panuse psühhiaatriliste häirete täieliku mõistmise saavutamiseks peetakse nüüd oluliseks mitme bioloogilise taseme hindamist, mida rahvasuus nimetatakse "oomika "lähenemiseks. 21 Joonisel 1 on kujutatud erinevad bioloogilised tasemed, kus saab hinnata kõiki viit süsteemi, ja potentsiaalsed markerite allikad, millele neid hinnanguid saab panna. Pange tähele, et kuigi iga süsteemi saab kontrollida igal oomikatasandil, on optimaalsed mõõteallikad igal tasandil selgelt erinevad. Näiteks pakub neuroimaging platvormi aju struktuuri või funktsiooni kaudseks hindamiseks, samas kui vere valguuuringud hindavad markereid otseselt. Transkriptoomika22 ja metaboloomika23 on üha populaarsemad, pakkudes hinnangut potentsiaalselt tohutule arvule markeritele. Inimese mikrobioomiprojekt üritab nüüd tuvastada kõiki inimese mikroorganisme ja nende geneetilist koostist. 24 Uudsed tehnoloogiad parandavad meie võimet neid mõõta, sealhulgas täiendavate allikate kaudu ; näiteks hormoone, näiteks kortisooli, saab nüüd analüüsida juustes või küüntel (pakkudes kroonilist näidustust) või higist (pakkudes pidevat mõõtmist), 25 aga ka veres, tserebrospinaalvedelikus, uriinis ja süljes.
Arvestades depressiooniga seotud oletatavate allikate, tasemete ja süsteemide arvu, pole üllatav, et translatsioonipotentsiaaliga biomarkerite skaala on ulatuslik. Eriti kui mõelda markerite vastastikmõjudele, on tõenäoliselt ebatõenäoline, et üksikute biomarkerite eraldi uurimine annab tulemusi kliinilise praktika parandamiseks. Schmidt jt soovitasid kasutada biomarkerite paneele ja seejärel esitasid Brand jt 26 visandipaneeli eelnõu, mis põhines varasematel MDD kliinilistel ja prekliinilistel tõenditel, tuvastades 27 "tugevat" biomarkeri sihtmärki, millest igaüks on harva üks marker. Need hõlmavad vähenenud halli aine mahtu (hipokampuses, prefrontaalses ajukoores ja basaalganglioni piirkondades), ööpäevase tsükli muutusi, hüperkortisolismi ja muid hüpotalamuse hüpofüüsi- neerupealise (HPA) telje hüperaktivatsiooni, kilpnäärme talitlushäireid, vähendatud dopamiini, noradrenaliini või 16-hüdroksüindooläädikhapet. , suurenenud glutamaadi sisaldus, suurenenud superoksiiddismutaasi ja lipiidide peroksüdatsioon, nõrgestatud tsüklilise adenosiini 5a, 3a-monofosfaadi ja mitogeeniga aktiveeritud valgukinaasi raja aktiivsus, suurenenud põletikuvastased tsütokiinid, muutused trüptofaanis, künureniinis, insuliinis ja spetsiifilised geneetilised polümorfismid. Need markerid ei ole konsensuse alusel kokku lepitud ja neid saab mõõta mitmel viisil; on selge, et keskendunud ja süstemaatiline töö peab selle tohutu ülesandega tegelema, et tõestada nende kliinilist kasu.
Selle läbivaatamise eesmärgid
Tahtlikult laiaulatusliku ülevaate saamiseks püütakse käesoleva artikliga määrata kindlaks biomarkerite uuringute üldised vajadused depressioonis ja seda, mil määral on biomarkeritel reaalne translatsioonipotentsiaal ravimite ravivastuse suurendamiseks. Alustame arutades selle valdkonna kõige olulisemaid ja põnevaid tulemusi ning suunates lugeja täpsematele ülevaatustele asjakohaste markerite ja võrdluste kohta. Me kirjeldame praeguseid tõendeid silmas pidades tekkivaid probleeme koos depressiooni koormuse vähendamise vajadustega. Lõpuks ootame olulisi uurimisvõimalusi praeguste väljakutsetega toimetulemiseks ja nende mõju kliinilisele praktikale.
Viimased Insights
Depressiooniga inimestele mõeldud kliiniliselt kasulike biomarkerite otsimine on toonud kaasa ulatusliku uurimise viimase poole sajandi jooksul. Kõige sagedamini kasutatavad ravimeetodid olid loodud monoamiini depressiooni teooriast; hiljem suurt tähelepanu pöörati neuroendokriinsetele hüpoteesidele. Viimastel aastatel on kõige viljakamad uuringud ümbritsetud depressiooni põletikulise hüpoteesi. Kuid paljud viited läbi viitavad artiklid on keskendunud kõigile viiele süsteemile; vt tabelit 1 ja allpool, et koguda hiljutisi teadmisi kogu biomarkeri süsteemide kohta. Paljudel tasemetel mõõdetuna on kõige sagedamini uuritud verest saadud valgud ja pakuvad biomarkerite allikaid, mis on mugav, kulutõhusad ja võivad olla translatsioonipotentsiaalist lähemal kui muud allikad; seega täpsemalt antakse vere ringlevatele biomarkeritele.
Hiljutises süstemaatilises ülevaates uurisid Jani jt20 perifeersel verel põhinevaid depressiooni biomarkereid koos ravitulemustega. Ainult 14 kaasatud uuringust (otsiti kuni 2013. aasta alguseni) uuriti 36 biomarkerit, millest 12 olid vähemalt ühe uurimise käigus olulised vaimse või füüsilise reageerimise indeksite ennustajad. Need, mis tuvastati potentsiaalselt mittevastamise riskifaktoritena, hõlmasid põletikulisi valke: madal interleukiin (IL) -12p70, lümfotsüütide ja monotsüütide arvu suhe; neuroendokriinsed markerid (kortisooli mittesummutav deksametasoon, kõrge vereringes kortisool, vähendatud kilpnääret stimuleeriv hormoon); neurotransmitteri markerid (madal serotoniini ja noradrenaliini sisaldus); metaboolsed (madala tihedusega lipoproteiinide kolesterool) ja neurotroofsed tegurid (vähendatud S100 kaltsiumi siduv valk B). Lisaks sellele on teistes ülevaadetes teatatud seostest täiendavate biomarkerite ja ravitulemuste vahel. 19,28, 30, 2 Iga süsteemi oletatavate markerite lühikirjeldus on esitatud järgmistes osades ja tabelis XNUMX.
Depressiooni põletikulised leiud
Alates Smithi makrofagihüpoteesi kirjeldavast põhitööst 31 on see väljakujunenud kirjandus leidnud depressiooniga patsientidel suurenenud erinevate põletikuvastaste põletikumarkerite tasemeid, mida on laialdaselt läbi vaadatud. kontrollivad populatsioone.32 37
IL-6 (P <0.001 kõigis metaanalüüsides; kaasa arvatud 31 uuringut) ja CRP (P <0.001; 20 uuringut) ilmnevad depressioonis sageli ja usaldusväärselt kõrgenenud. Varasetes uuringutes tuvastati alfa kasvaja nekroosifaktori (TNFa) tõus. (P <40), 0.001, kuid oluline heterogeensus muutis selle hiljutiste uuringute arvestamisel (38 uuringut) veenvaks. 31 IL-40? on depressiooniga veelgi ebaselgemalt seotud, metaanalüüsid viitavad depressiooni kõrgemale tasemele (P = 1), ainult Euroopa uuringutes 0.03 kõrgele tasemele41 või kontrollide erinevuste puudumisele. 42 Vaatamata sellele soovitas hiljutine artikkel IL-le erilisi translatiivseid tagajärgi 40a, 1, mida toetab kõrgenenud IL-44a ülimalt oluline mõju ribonukleiinhape, mis ennustab halba vastust antidepressantidele; 1 muud ülaltoodud leiutist on seotud vereringes verest saadud tsütokiinidega. Kemokiinmonotsüütide kemoattraktantvalk-45 on ühe metaanalüüsi käigus näidanud depressioonis osalejate tõusu.1 Interleukiinid IL-39, IL-2, IL-4, IL-8 ja gammainterferoon ei erinenud depressiooniga patsientide ja metaanalüütilisel tasemel, kuid on siiski näidanud ravi muutmise potentsiaali: on teatatud, et IL-10 on prospektiivselt ja ristlõikeliselt raskekujulise depressiooniga inimestel kõrgenenud, IL-8 ja gamma-interferooni muutuste 46 erinevat mustrit ravi ajal on ilmnenud varajaste ravivastajate ja mittevastajate vahel, 10, samas kui IL-47 ja IL-4 on vähenenud vastavalt sümptomite remissioonile.2 Metaanalüüsides on IL-48, IL-6a, IL- 1 ja CRP.10. Lisaks on TNFa? võib ravivastuses ravivastuses väheneda ja kombineeritud markerindeks võib viidata suurenenud põletikule patsientidel, kes hiljem ravile ei reageeri. 43,49,50 On siiski märkimisväärne, et peaaegu kõigis põletikulisi valke ja ravivastust uurivates uuringutes kasutatakse farmakoloogilisi ravikatseid . Seega on vähemalt mõned põletikulised muutused ravi ajal tõenäoliselt seotud antidepressantidega. Erinevate antidepressantide täpseid põletikulisi toimeid pole veel kindlaks tehtud, kuid CRP tasemeid kasutavate tõendite kohaselt võivad inimesed reageerida spetsiifilisele ravile lähtuvalt algtaseme põletikust: Harley jt43 teatasid kõrgenenud CRP-eelsest ravist, ennustades halba vastust psühholoogilisele ravile (kognitiivne, käitumuslik või inimestevaheline psühhoteraapia), kuid hea reaktsioon nortriptüliinile või fluoksetiinile; Uher jt 51 kordasid seda leidu nortriptüliini suhtes ja tuvastasid estsitalopraami vastupidise efekti. Seevastu leidsid Chang jt52 fluoksetiinile või venlafaksiinile varajase raviga reageerijatest suurema CRP kui mittevastavad. Lisaks on TRD ja kõrge CRP-ga patsiendid paremini reageerinud TNF? infliksimabi antagonist kui normaalses vahemikus
Koos näitavad tõendid, et isegi kui kontrollida selliseid tegureid nagu kehamassiindeks (BMI) ja vanus, võivad põletikulised reaktsioonid olla ebaregulaarsed ligikaudu kolmandikul depressiooniga patsientidest. 55,56 põletikuline süsteem on siiski väga keeruline ja on olemas arvukad biomarkerid, mis esindavad selle süsteemi erinevaid aspekte. Hiljuti on täiendavad uudsed tsütokiinid ja kemokiinid andnud tõendeid depressiooni kõrvalekallete kohta. Nende hulka kuuluvad: makrofaagide pärssiv valk 1a, IL-1a, IL-7, IL-12p70, IL-13, IL-15 eotaksiin, granulotsüütide-makrofaagide kolooniaid stimuleeriv faktor, 57 IL-5,58 IL-16,59 IL-17,60 monotsüütide kemoatraktantvalk -4,61 harknääre ja aktivatsiooniga reguleeritavat kemokiinist 62 eotaksiiniga 3 TNFb, 63 interferoongammast indutseeritud valgu 10,64 seerumi amüloid-A 65 lahustuva rakusisese haardumisega molecule66 ja lahustuva veresoonte rakuadhesioonimolekul 1.67
Kasvufaktori leiud depressioonis
Pidades silmas mitte-neurotroofsete kasvufaktorite (nagu need, mis on seotud angiogeneesiga) potentsiaalse tähtsusega, viidatakse neurogeensetele biomarkeritele kasvufaktorite laiema määratluse all.
Neist kõige sagedamini uuritakse ajust pärinevat neurotroofset tegurit (BDNF). Mitmed metaanalüüsid demonstreerivad seerumi BDNF-valgu nõrgenemist, mis näib suurenevat koos antidepressantidega. 68. Viimane neist analüüsidest viitab sellele, et need BDNF-i kõrvalekalded on rohkem väljendunud kõige raskemini depressiooniga patsientidel, kuid antidepressandid näivad olevat suurendavad selle valgu taset isegi kliinilise remissiooni puudumisel. 71 proBDNF-i on uuritud vähem kui BDNF-i küpset vormi, kuid need kaks näivad funktsionaalselt (mõju poolest türosiiniretseptori kinaas-B retseptoritele) ja hiljutised tõendid näitavad, et kuigi küps BDNF võib depressioonis väheneda, võib proBDNF olla üleprodukteeritud. 70 Perifeerselt hinnatud närvikasvufaktorit on metaanalüüsis täheldatud ka madalamas depressioonis kui kontrollis, kuid antidepressantravi ei pruugi seda muuta, hoolimata sellest, et kõige nõrgem raskema depressiooniga patsientidel.72 Gliarakkude metaanalüüsis on kirjeldatud sarnaseid leideliinist tuletatud neurotroofne tegur.73
Vaskulaarne endoteeli kasvufaktor (VEGF) mängib koos teiste VEGF-i perekonna liikmetega (nt VEGF-C, VEGF-D) angiogeneesi ja neurogeneesi edendamisel rolli ning lubab depressiooni. 75 Vaatamata ebajärjekindlatele tõenditele on kaks metaanalüüsi hiljuti näidatud VEGF-i tõus depressiooniga patsientide veres võrreldes kontrollrühmadega (16 uuringus; P <0.001). 76,77 Siiski on TRD78-s tuvastatud madal VEGF ja kõrgemad tasemed on ennustanud antidepressantidega mittevastamist.79 Ei ole mõistetav miks VEGF-valgu tase tõuseb, kuid osaliselt võib see olla tingitud põletikueelsest aktiivsusest ja / või vere aju barjääri läbilaskvuse suurenemisest depressioonis, mis põhjustab vähenenud ekspressiooni tserebrospinaalvedelikus.80 VEGF-i ja ravivastuse suhe on ebaselge ; hiljutises uuringus ei leitud seoseid seerumi VEGF-i ega BDNF-i vahel ravivastuse või depressiooni raskusastmega, vaatamata antidepressantravi vähenemisele.81 Insuliinitaoline kasvufaktor-1 on neurogeensete funktsioonidega täiendav tegur, mis võib depressioonis suureneda, peegeldades tasakaaluhäireid neurotroofsed protsessid.82,83 Põhiline fibroblastide kasvufaktor (või FGF-2) on fibroblastide kasvufaktorite perekonna liige ja tundub kõrgem depressioonis kui kontrollrühmades.84 Kuid aruanded pole järjepidevad; üks leidis, et see valk oli MDD-s madalam kui tervislikel kontrollidel, kuid vähenes antidepressantravi kõrval veelgi
Edasine kasvufaktoreid, mida ei ole piisavalt uuritud Depressiooni hulka türosiinkinaasi 2 ja lahustuva fms-sarnane türosiinkinaas-1 (nimetatud ka sVEGFR-1), mis toimivad sünergias VEGF ja türosiini kinaasi retseptorite (mis seostuvad BDNF) võib nõrgestatud depressioonis. 86 Platsentaarse kasvufaktor on ka osa VEGF-i perekonnast, kuid meie teadmistest ei ole süstemaatiliselt allasurutud proovides uuritud.
Metaboolsed biomarkerid depressiooni kohta
Peamiseks biomarkerite seostatud metaboolsete haiguse hulka leptiini adiponektiinil greliini, triglütseriidide kõrge tihedusega lipoproteiini (HDL), glükoos, insuliin ja albumin.87 omavahelised seosed paljud neist ja depressioon on üle vaadatud: leptin88 ja ghrelin89 ilmuvad madalam depressiooni kui kontrollid perifeersetes piirkondades ja võivad suureneda koos antidepressantidega või remissiooniga. Depressioonis võib insuliiniresistentsust suurendada, ehkki väikestes kogustes. 90 Lipiidiprofiilid, sealhulgas HDL-kolesterool, ilmnevad paljudel depressiooniga patsientidel, sealhulgas depressioonitajatega kaasnevate füüsiliste haiguste korral, kuid see suhe on keeruline ja vajab täiendavat selgitamist. 91 Lisaks sellele, ülevaates on teatatud depressioonist põhjustatud hüperglükeemiat92 ja hüpoalbuminaemia93.
Üldiste metaboolsete seisundite uurimine muutub üha sagedasemaks, kasutades väikeste molekulide metaboloomikapaneele, lootuses leida psühhiaatriliste häirete korral tugev biokeemiline allkiri. Hiljutises tehisintellekti modelleerimist käsitlevas uuringus ennustas metaboliitide komplekt, mis illustreerib suurenenud glükoosi lipiidide signaalimist, MDD diagnoosi, mis toetas varasemaid uuringuid.
Neurotransmitteri leiud depressioonis
Ehkki depressioonis monoamiinidele pööratud tähelepanu on andnud suhteliselt edukad ravimeetodid, pole antidepressantide monoamiinmärkide selektiivsuse põhjal ravi optimeerimiseks kindlaks tehtud ühtegi tugevat neurotransmittermarkerit. Hiljutised tööpõhimõtted serotoniini (5-hüdroksütrüptamiin) 1A retseptori poole on potentsiaalselt olulised nii depressiooni diagnoosimisel kui ka prognoosimisel, kuni oodatakse uusi geneetilisi ja pildistamismeetodeid.96 On uusi potentsiaalseid ravimeetodeid, mis on suunatud 5-hüdroksütrüptamiinile; näiteks 5-hüdroksütrüptofaani aeglase vabanemisega manustamine. 97 Dopamiini suurenenud levik interakteerub teiste neurotransmitteritega, et parandada kognitiivseid tulemusi, näiteks otsuste langetamist ja motivatsiooni. depressiooniga seotud stressivastuse osana; see võib üleujutuse kaudu vähendada 98-hüdroksütrüptamiini tootmist. Hiljutises ülevaates esitatakse see teooria ja soovitatakse, et TRD-s võiks seda muuta (ja 5-HT taastada) multimodaalse ravi abil, mis on suunatud mitmele neurotransmitterile. 5 Huvitaval kombel ei esine serotoniini tõus alati koos terapeutiliste antidepressantide eeliste on leitud, et neurotransmitterite metaboliidid, näiteks 99-metoksü-100-hüdroksüfenüülglükool, noradrenaliin või dopamiini homovanilliinhape, suurenevad depressiooni vähendamise kõrval antidepressantidega 3 või et nende metaboliitide madal tase ennustab paremat vastust SSRI ravi. 4
Neuroendokriinne depressiooni leiud
Kortisool on kõige levinum HPA telje biomarker, mida on uuritud depressioonis. Arvukad arvustused on keskendunud HPA-tegevuse erinevatele hinnangutele; üldiselt viitavad sellele, et depressioon on seotud hüperkortisoleemiaga ja et kortisooli äratõukereaktsioon on sageli nõrgestatud. 104,105 Seda toetab hiljuti läbi viidud krooniliste kortisooli tasemete mõõtmine juustes, toetades kortisooli hüperaktiivsuse hüpoteesi depressioonis, kuid hüpoaktiivsus muudes haigustes nagu paanikahäire. 106 Lisaks võib eriti kortisooli kõrgenenud taset ennustada vaesemat vastust psühholoogilisele107 ja antidepressandi 108 ravile. Ajalooliselt on perspektiivse ravivastuse kõige levinum neuroendokriinsed markerid deksametasooni supresseerimiskatse, kus pärast deksametasooni manustamist seostatakse kortisooli mitte-supresseerimist järgneva remissiooni väiksema tõenäosusega. Kuid seda nähtust ei peetud kliiniliseks rakendamiseks piisavalt tugevaks. Seostuvad markerid - kortikotropiini vabastavat hormooni ja adrenokortikotropiini hormooni ning vasopressiini - leiti olevat ebajärjekindel kui depressioonis ületootmine ja leitakse, et dehüdroepiandrosteroon on nõrgestatud; suhe kortisooli dehüdroepiandresterooniga võib suureneda ka suhteliselt stabiilsed SM TRD, kestis pärast remission.109 Neuroendokriinsed hormooni düsfunktsioonid juba ammu seostatud depressiooni, ja hüpotüreoidism võib samuti mängida määravat rolli depressiivsetel mood.110 Lisaks kilpnäärme vastuseid saab depressiooni edukaks raviks normaliseerida. 111
Ülaltoodu raames on oluline arvestada ka signaaliradadega süsteemides, näiteks glükogeeni süntaasi kinaas-3, mitogeeniga aktiveeritud valgukinaas ja tsükliline adenosiin 3a, 5a-monofosfaat, mis on seotud sünaptilise plastilisusega112 ja modifitseeritud antidepressantidega. potentsiaalseid biomarkerite kandidaate, mis hõlmavad eriti bioloogilisi süsteeme, mõõdetakse neurokujutiste või geneetika abil. Reageerides depressiivsete ja masendumata populatsioonide vaheliste tugevate ja sisuliste genoomiliste erinevuste puudumisele, võiks 113 uut geneetilist lähenemisviisi, näiteks polügeense skoori 114 või telomeeri pikkuse 115 116,117, osutuda kasulikumaks. Täiendavad populaarsust koguvad biomarkerid uurivad ööpäevaseid tsükleid või kronobioloogilisi biomarkereid, kasutades erinevaid allikaid. Aktigraafia abil saab kiirendusmõõturi abil objektiivselt hinnata une ja ärkveloleku aktiivsust ning puhkeaega ning aktigraafilised seadmed saavad üha enam mõõta lisategureid, näiteks valguse kiirgust. See võib olla avastamiseks kasulikum kui patsientide üldkasutatavad subjektiivsed aruanded ja pakkuda ravivastuse uudseid ennustajaid.118 Küsimus, millised biomarkerid on translatsioonikasutuses kõige lootustandvamad, on keeruline, mida laiendatakse allpool.
Praegused väljakutsed
Nende viie vaatlusaluse neurobioloogilise süsteemi kohta on tõendeid järgnenud sarnane narratiiv: on olemas palju biomarkereid, mis on mõnes mõttes seotud depressiooniga. Need markerid on tihti omavahel seotud kompleksselt, raskesti modereeritud viisil. Tõendid on ebajärjekindlad ja on tõenäoline, et mõni neist on teiste tegurite epifenomena, millest mõned on olulised ainult mõnedes patsientidel. Biomarkerid on tõenäoliselt kasulikud erinevatel viisidel (nt need, mis ennustavad järgnevat ravivastust, need, mis näitavad, et spetsiifilised ravimeetodid on tõenäolisemalt efektiivsed või need, mis muudavad sekkumist kliiniliste paranemisest hoolimata). Bioloogiliste hindamiste järjepidevuse ja kliinilise rakendatavuse maksimeerimiseks psühhiaatriapopulatsioonides on vaja uudseid meetodeid.
Biomarkeri varieeruvus
Biomarkerite muutused aja jooksul ja erinevates olukordades mõjutavad teatud tüüpi (nt proteoomika) rohkem kui teised (genoomika). Paljudele standardsetele normidele pole olemas või neid ei ole laialdaselt aktsepteeritud. Tõepoolest, keskkonnategurite mõju markeritele sõltub tihti geneetilistest koostistest ja muudest füsioloogilistest erinevustest inimestele, keda ei saa kõiki arvesse võtta. See muudab biomarkeri aktiivsuse hindamise ja bioloogiliste kõrvalekallete kindlakstegemise, mida on raske tõlgendada. Potentsiaalsete biomarkerite arvu tõttu pole paljusid teisi asjakohaseid markereid kasutades laialdaselt mõõdetud või kogu paneel.
On teatatud paljudest teguritest, mis muudavad afektiivsete häiretega patsientidel valkude taset bioloogilistes süsteemides. Nende hulka kuuluvad teadusuuringutega seotud tegurid, näiteks kestus ja säilitamistingimused (mis võivad põhjustada mõne ühendi halvenemist), nende hulka kuuluvad mõõdetud päevade arv, rahvus, kehaline koormus, 119-toitumine (nt mikrobiomia aktiivsus, eriti tingimusel, et enamik vere biomarkeri uuringuid teevad 120 suitsetamine ja ainete kasutamine, 121, samuti tervisetegurid (nt kaasuvate põletikuliste, kardiovaskulaarsete või muude füüsiliste haiguste korral). Näiteks, kuigi depressiooniga, kuid teisiti tervislike indiviidide puhul on depressioonis olevatel rühmadel kõrgendatud põletikku täheldatud, on depressioonis inimestel, kellel on ka kaasuva immuunsusega seotud haigusseisund, sagedamini tsütokiinide tase isegi kõrgem kui neil, kellel puudub depressioon või haigus. 122 Mõned silmapaistvad faktorid biomarkerite seos, depressioon ja ravivastus on tõenäoliselt kaasatud allpool.
Stress. Mõlemad endokriinsed ja immuunvastused omavad stressi (füsioloogilist või psühholoogilist) reageerimiseks hästi tuntud rolli ning uurimisuuringutes mõõdetakse bioloogiliste isendite kogumise ajal mööduvat stressi, hoolimata selle faktori varieeruvusest üksikisikute vahel, mida praegused depressiivsed sümptomid. Mõlemad ägedad ja kroonilised psühholoogilised stressorid toimivad immuunprobleemina, rõhutades põletikulisi reaktsioone lühikeses ja pikemas perspektiivis. 123,124 See avastus laieneb varajase stressi tekkele, mida on seostatud täiskasvanute põletikuliste tõusudega, mis on sõltumatud stressist täiskasvanu. 125,126 Lapseea traumaatilisel kogemusel on täheldatud ka kõrgendatud põletikku ainult nendel lastel, kes olid hetkel depressioonis. 127 Seevastu depressiooni ja lapseea trauma ajaloos olevatel inimestel võib olla pingest tingitud kortisooli vastus pinge all, võrreldes depressiooni ja puudub varajase elu trauma.128 Stress-indutseeritud HPA-telje muutused on omavahel seotud kognitiivse funktsiooniga, 129-ga, samuti depressiooni alatüübiga või HPA-ga seotud geenide varieerumisega. 130 Stressil on ka lühi- ja pikaajaline kahjustus neurogeneesile131 ja teiste neuronite mehhanismid. 132 Täpselt on selge, kuidas lapsepõlve trauma mõjutab bioloogilisi markereid surudes ed täiskasvanutel, kuid on võimalik, et varajase eluea stress paneb mõnedel inimestel täiskasvanueas püsivaid stressireaktsioone, mida psühholoogiliselt ja / või bioloogiliselt võimendatakse.
Kognitiivne toimimine. Neurokognitiivsed häired esinevad sageli inimestel, kellel on afektiivsed häired, isegi ravimata MDD-is. 133 Kognitiivsed puudujäägid ilmnevad kumulatiivselt koos ravitakistusega. 134 Neurobioloogiliselt on sellel suhetel võtmeroll HPA teljel 129 ja neurotroofsed süsteemid 135. Neurotransmitterid noradrenaliin ja dopamiin on tõenäoliselt olulised kognitiivsete protsesside jaoks, nagu õppimine ja mälu. 136 Kõrgendatud põletikulised reaktsioonid on seotud kognitiivse langusega ja võivad mõjutada kognitiivset funktsiooni depressiooni episoodides, 137-il ja remissioonil erinevate mehhanismide kaudu. 138 Tõepoolest, Krogh ja al139 pakkusid, et CRP on paremini seotud kognitiivse toimega kui depressiooni sümptomitega.
Vanus, sugu ja kehamassiindeks. Meeste ja naiste bioloogiliste erinevuste puudumine või esinemine ja nende suundumus on senini tõestatud eriti varieeruv. Neuroendokriinhormooni varieeruvus meeste ja naiste vahel mõjutab depressiooni tundlikkust. 140 Põletiku uuringute ülevaates on teatatud, et vanuse ja soo kontrollimine ei mõjutanud põletikuliste tsütokiinide patsientide kontrolli erinevusi (kuigi IL-6 ja depressiooni seos vähenes vanuse suurenedes, mis on kooskõlas teooriatega, mille kohaselt suureneb tavaliselt põletik koos vanusega). 41,141 VEGF-i erinevused patsientide ja kontrollide vahel on nooremate proovide hindamisel uuringutes suuremad, samas kui sugu, KMI ja kliinilised tegurid ei mõjuta neid võrdlusi metaanalüütilisel tasemel. 77 Kuid BMI-de kohandamise puudumine varasemates põletiku- ja depressiooniuuringutes näib segavat nende rühmade vahel esinenud väga olulisi erinevusi. 41 Laiendatud rasvkoe on näidanud kindlalt, et stimuleerib tsütokiinide tootmist ja on tihedalt seotud metaboolsete markeritega. 142 Kuna psühhotroopsed ravimid võib olla seotud kellel on kehakaalu tõus ja suurem BMI, ning neid on seostatud ravivastusega depressioonis, on see oluline uurimisvaldkond.
Ravimeid. Paljud biomarkerite uuringud depressioonis (nii ristlõike- kui ka pikisuunalised) on heterogeensuse vähendamiseks kogunud ravimpreparaatidelt lähteproovid. Kuid paljud neist hinnangutest võetakse pärast ravimite väljapesu perioodi, mis jätab potentsiaalselt märkimisväärse segava faktori järelejäänud muutused füsioloogias, mida võimendab lai valik ravimeetodeid, millel võib olla erinev mõju põletikule. Mõnes uuringus on psühhotroopne, kuid mitte muude ravimite kasutamine välistatud: eriti on suukaudsed rasestumisvastased tabletid uuringus osalejatel sageli lubatud ja seda ei saa analüüsides kontrollida, mis on hiljuti näidanud hormoonide ja tsütokiinide taseme tõstmist. 143,144, 34,43,49,145 Mitmed uuringud näitavad, et antidepressandid ravimitel on mõju põletikulisele reaktsioonile, 147 HPA-teljele, 108 neurotransmitterile, 148 ja neurotroofsele 149 aktiivsusele. Kuid arvukatel potentsiaalsetel depressiooni ravimeetoditel on selged ja keerulised farmakoloogilised omadused, mis viitab sellele, et praeguste andmete põhjal võib erinevate ravivõimaluste diskreetne bioloogiline toime olla. On väidetud, et lisaks monoamiinimõjudele suunavad spetsiifilised serotoniini sihtivad ravimid (st SSRI-d) tõenäoliselt ka põletiku Th2 nihkeid ja noradrenergilised antidepressandid (nt SNRI-d) Th1-nihet.150 Ei ole veel võimalik määrata üksikute või kombineeritud ravimite mõju biomarkeritele. Neid vahendavad tõenäoliselt muud tegurid, sealhulgas ravi pikkus (vähestes uuringutes hinnatakse pikaajalist ravimite kasutamist), valimi heterogeensus ja osalejate mitte stratifitseerimine ravivastusega.
Heterogeensus
Metoodiline. Nagu ülalpool mainitud, on erinevused (uuringute ajal ja sees), mille kohaselt osalejad saavad ja on eelnevalt võtnud ravimeetodid (ja kombinatsioonid), seostada uurimistulemuste heterogeensusega, eriti biomarkerite uuringutega. Lisaks sellele erinevad uuringud ka paljudest muudest projekteerimis- ja näidisobjektide omadustest, mis suurendavad tõlgendamise raskusi ja tulemuste omistamist. Nende hulka kuuluvad biomarkeri mõõtmisparameetrid (nt testimiskomplektid) ja depressiooni markerite kogumise, säilitamise, töötlemise ja analüüsimise meetodid. Hiles et al141 uuris põletikuvastases kirjanduses mitmesuguseid vasturääkivuse allikaid ning leidsid, et depressiooni ja depressiooniga rühmade perifeerse põletiku hindamisel on kõige olulisem depressiooni diagnoosimise, BMI ja kaasuvate haiguste täpsus.
Kliiniline. Depresseeritud populatsioonide ulatuslik heterogeensus on hästi dokumenteeritud151-is ja on kriitiline kaasaja uurimistöö kirjanduses olevate kontrastsete leidude jaoks. On tõenäoline, et isegi diagnooside ajal võivad ebanormaalsed bioloogilised profiilid piirduda üksikisikute alamhulkadega, mis ei pruugi aja jooksul olla stabiilsed. Depressiooniga inimeste koherentsed alarühmad võivad olla identifitseeritavad psühholoogiliste ja bioloogiliste tegurite kombinatsiooni kaudu. Allpool kirjeldame potentsiaali alarühmade uurimiseks biomarkeri varieeruvuse ja heterogeensuse probleemide lahendamisel.
Depressiooni alltüübid
Siiani pole ükski depressiooniepisoodide või häirete homogeenne alarühm suutnud usaldusväärselt eristada patsiente sümptomite esitusviisi või ravivastuse põhjal. 152 Alarühma olemasolu, kus bioloogilised kõrvalekalded on rohkem väljendunud, aitaks selgitada varasemate uuringute heterogeensust. võiks katalüüsida kihistunud ravi suunas. Kunugi jt. 153 on pakkunud nelja võimaliku alamtüübi komplekti, mis põhinevad erinevate neurobioloogiliste süsteemide rollil, millel on depressioonis kliiniliselt olulised alatüübid: hüperkortisolismiga melanhoolne depressioon või hüpokortisolism, mis peegeldab ebatüüpilist alamtüüpi, dopamiiniga seotud patsientide alamhulka esinevad silmapaistvalt anhedooniaga (ja võivad hästi reageerida näiteks aripiprasoolile) ja põletikulise alatüübiga, mida iseloomustab kõrgenenud põletik. Paljud põletikule keskenduvad artiklid on täpsustanud põletikulise alatüübi olemasolu depressioonis. 55,56,154,155 Kõrgenenud põletiku kliinilised korrelaadid pole veel määratletud ja väheseid otseseid katseid on tehtud selle kohordi koosseisu avastamiseks. On välja pakutud, et ebatüüpilise depressiooniga inimestel võib olla kõrgem põletiku tase kui melanhoolse alatüübi korral 156, mis pole võib-olla kooskõlas järeldustega, mis käsitlevad HPA telge melanhoolse ja ebatüüpilise depressiooni alatüübi korral. TRD37 või silmapaistvate somaatiliste sümptomitega depressioon157 on samuti viidatud potentsiaalsele põletikulisele alatüübile, kuid neurovegetatiivsed (uni, söögiisu, libiido kadu), meeleolu (sealhulgas madal meeleolu, suitsidaalsus ja ärrituvus) ja kognitiivsed sümptomid (sealhulgas afektiivne kallutatus ja süütunne) 158 näivad olevat seotud bioloogiliste profiilidega. Põletikulise alatüübi potentsiaalsed kandidaadid hõlmavad haigusekäitumisega sarnaseid sümptomeid159,160 158 või metaboolset sündroomi. XNUMX
Kalduvus (hüpo-maania) suunas võib bioloogiliselt eristada depressiooni all kannatavate patsientide vahel. Tõendid nüüd viitavad sellele, et bipolaarsed haigused on mitmekülgsed meeleoluhäirete rühmad, mille bipolaarne subitsioosne häire leiti rohkem levinud kui varem tunnustatud. 161 Hiljuti on kliinilise psühhiaatria peamine probleem esile tõstetud bipolaarse häire ebatäpsuse ja / või hilinemisega, kusjuures keskmine aeg diagnoosi korrigeerimiseks, mis sageli ületab kümnendit162i ja see viivitus põhjustab üldist haigust suuremat raskust ja maksumust. 163 Enamikul bipolaarse häirega patsientidest, kes esialgu esinesid ühe või enama depressiivse episoodiga ja kõige sagedasem vale diagnoosimine oli unipolaarse depressiooni korral, tegurid, mis võivad eristada unipolaarset ja bipolaarse depressiooni, omavad olulist mõju. 164 Bipolaarsete spektrihäirete tõenäosus on mõnede varasemate MDD biomarkerite uuringutes tõenäoliselt avastamata ja tõendeid peegeldades näidanud HPA-telje aktiivsuse 109 või põletiku165,166 diferentseerumist bipolaarse ja unipo vahel lar depressioon. Siiski on need võrdlused nappid, neil on väikeste valimite suurused, tuvastatud märkimisväärsed trendi mõjud või värbatud populatsioonid, mida ei diagnoositud hästi. Nendes uuringutes ei uurita ka ravivastuse rolli nendes suhetes.
Mõlemad bipolaarsed häired167 ja ravimiresistentsus168 ei ole dikotoomilised konstruktsioonid ja paiknevad pidevalt, mis suurendab alatüübi tuvastamist. Lisaks alamtüübile väärib märkimist, et paljud depressiooniga täheldatud bioloogilised kõrvalekalded leiavad sarnaselt teiste diagnoosidega patsientidel. Seega on transdiagnostilised uuringud ka potentsiaalselt olulised.
Biomarkeri mõõtmise väljakutsed
Biomarkeri valik. Suur hulk potentsiaalselt kasulikke biomarkereid esitab psühhobioloogiale väljakutse, et teha kindlaks, millised markerid millisel viisil ja kelle jaoks on seotud. Väljakutse suurendamiseks on suhteliselt vähe neist biomarkeritest depressiooni osas piisavalt uuritud ja enamiku jaoks pole nende täpne roll tervetes ja kliinilistes populatsioonides hästi teada. Vaatamata sellele on mitmeid katseid välja pakkuda paljutõotavaid biomarkeripaneele. Lisaks Brandi jt 16 tugeva potentsiaaliga markerite komplektile kirjeldavad 27 Lopresti jt täiendavat ulatuslikku oksüdatiivse stressi markerite komplekti, millel on potentsiaal ravivastuse parandamiseks. 28 Papakostas jt määratlesid a priori üheksast seerumimarkerist koosneva komplekti bioloogilised süsteemid (BDNF, kortisool, lahustuv II tüüpi TNFa retseptor, alfa1 antitrüpsiin, apolipoproteiin CIII, epidermise kasvufaktor, müeloperoksidaas, prolaktiin ja resistiin) valideerimis- ja replikatsiooniproovides MDD-ga. Kui need ühendati, suutis nende tasemete liitmõõt eristada MDD ja kontrollrühmi 80–90% täpsusega.169. Me soovitame, et isegi need ei hõlmaks kõiki selle valdkonna potentsiaalseid kandidaate; vt tabel 2 depressioonipotentsiaaliga biomarkerite ammendamatu piiritlemise kohta, mis sisaldab nii tõendusbaasi kui ka paljutõotavaid uudseid markereid.
Technology. Tehnoloogiliste edusammude tõttu on nüüd võimalik (tõepoolest, mugav) mõõta suure hulga biomarkerite samaaegselt madalama hinnaga ja suurema tundlikkusega kui varem. Praegu on see suutlikkus arvutada arvukaid ühendeid ennekõike meie võimet tõhusalt analüüsida ja tõlgendada andmeid, 170 midagi, mis jätkub biomarkeri massiivide kasvu ja uute markeritega, nagu näiteks metabomüütikaga. See on tingitud peamiselt sellest, et markerite täpne roll ja seosed ei ole arusaadavad, ning ebapiisav arusaam sellest, kuidas seostatud markerid seostuvad erinevate bioloogiliste tasemetega (nt geneetiline, transkriptsioon, proteiin) üksikisikute sees ja nende vahel. Uute analüütiliste lähenemisviiside ja standardite abil saadud suured andmed aitavad seda lahendada ja pakutakse uusi metoodikaid; Üks näide on statistilise lähenemisviisi väljatöötamine, mis põhineb voogu analüüsil, et avastada uusi potentsiaalseid metaboolseid markereid, mis põhinevad võrkudevahelisel reaktsioonil ja integreerivad geeniekspressiooni koos metaboliidi andmetega. 171 Masinõlkeid on juba rakendatud ja need aitavad kaasa biomarkerit kasutavatele mudelitele andmed suurte andmetega uuringute tulemuste prognoosimiseks. 172
Biomarkerite koondamine. Biomarkerite massiivi samaaegne uurimine on alternatiiv isoleeritud markerite kontrollimisele, mis võiks pakkuda täpsemat vaatepunkti bioloogiliste süsteemide või võrkude keerulisse võrku. 26 Samuti, et aidata selles kirjanduses siiani vastandlikke tõendeid lahti ühendada (eriti kui ja vastastikmõjud on hästi mõistetavad), siis saab biomarkerite andmeid liita või indekseerida. Üks väljakutse on selle teostamiseks optimaalse meetodi kindlakstegemine ja see võib vajada tehnoloogia täiustamist ja / või uudseid analüüsimeetodeid (vt jaotist Suured andmed). Ajalooliselt on kahe erineva biomarkeri vaheline suhe andnud huvitavaid tulemusi. 109,173 174 Biomarkerite andmeid on võimalik koondada suuremas mahus, näiteks neid, mis kasutavad põletikueelsete tsütokiinivõrkude põhikomponentide analüüsi.43 Metaanalüüsis on põletikueelsed tsütokiinid teisendati iga uuringu puhul ühekordse toimega suuruse skooriks ja üldiselt ilmnes enne antidepressantravi oluliselt kõrgem põletik, ennustades järgnevat mittevastamist ambulatoorsetes uuringutes. Komposiitbiomarkerite paneelid on nii väljakutse kui ka võimalus tulevaste uuringute jaoks, et teha kindlaks sisukad ja usaldusväärsed leiud, mida saab ravitulemuste parandamiseks kasutada. 80 Papakostase jt uuringus kasutati alternatiivset lähenemist, valides seerumi heterogeensete (põletikuliste, HPA telg ja metaboolsed süsteemid), mis olid eelmises uuringus näidatud depressioonis olevate ja kontrollgrupi isikute vahel erinevaks ning ühendasid need riskiskooriks, mis erines kahe sõltumatu valimi ning kontrollrühma tundlikkuse ja spetsiifilisusega> 169%.
Suured andmed. Suurandmete kasutamine on tõenäoliselt vajalik praeguste väljakutsetega tegelemiseks, mis on seotud heterogeensuse, biomarkerite varieeruvusega, optimaalsete markerite kindlakstegemiseks ja valdkonna suunamiseks depressiooni translatiivsete, rakendusuuringute poole. Kuid nagu eespool kirjeldatud, toob see kaasa tehnoloogilisi ja teaduslikke väljakutseid.175 Terviseteadused on hakanud suurandmete analüüsi kasutama alles hiljuti, kümmekond aastat hiljem kui ärisektoris. Kuid sellised uuringud nagu iSPOT-D152 ja konsortsiumid, näiteks Psühhiaatrilise geneetika konsortsium176, arenevad koos psühhiaatria bioloogiliste mehhanismide mõistmisega. Masinaõppimise algoritme on depressiooni biomarkerite puhul hakatud kasutama väga vähestes uuringutes: hiljutine uurimine koondas 5,000 biomarkeri> 250 osaleja andmeid; pärast andmete mitmekordset sisestamist viidi läbi masinõppega regressioon, mis näitas 21 potentsiaalset biomarkerit. Pärast täiendavaid regressioonianalüüse valiti välja kolm biomarkerit, mis seovad kõige tugevamalt depressiivsete sümptomitega (punaste vereliblede väga erinev suurus, seerumi glükoosi- ja bilirubiinisisaldus). Autorid järeldavad, et suuri andmeid saab hüpoteeside genereerimiseks tõhusalt kasutada. 177 Praegu on käimas suuremad biomarkerite fenotüüpide loomise projektid, mis aitavad meie depressiooni neurobioloogia tulevikku edasi viia.
Tuleviku väljavaated
Biomarkeri paneeli identifitseerimine
Kirjanduse senised leiud vajavad kordamist suuremahulistes uuringutes. See kehtib eriti uudsete biomarkerite kohta, nagu kemokiini harknääre ja aktivatsiooniga reguleeritud kemokiin ning kasvufaktor türosiinikinaas 2, mida meie teada pole kliiniliselt depressiooniga ja tervete kontrollproovidega uuritud. Suurandmete uuringud peavad analüüsima ulatuslikke biomarkerite paneele ja kasutama keerukaid analüüsimeetodeid, et täielikult kindlaks teha markerite seosed nende teguritega, mis neid kliinilises ja mittekliinilises populatsioonis muudavad. Lisaks võivad põhikomponentide analüüsi laiaulatuslikud kordused luua väga korreleerunud biomarkerite rühmad ja teavitada ka "komposiitide" kasutamist bioloogilises psühhiaatrias, mis võib suurendada tulevaste leidude homogeensust.
Homogeensete alamtüüpide avastamine
Biomarkeri valimise puhul võib olla vajalik mitu paneeli erinevate potentsiaalsete võimaluste jaoks, mida uuringud võiksid kaasa tuua. Kokkuvõttes näitavad praegused tõendid, et biomarkeriprofiilid on kindlasti, kuid kahtlemata muutunud depressiooni all kannatavate inimeste alapopulatsioonis. See võib olla diagnoosikategooriate sees või selle sees, mis võiks seletada mõne ebajärjekindluse tulemusi, mida käesolevas kirjanduses võib täheldada. Bioloogilise alamrühma (või alarühmade) kvantifitseerimist võib kõige paremini hõlbustada biomarkerivõrgu paneelide suurte klastrite analüüs depressioonis. See illustreeriks rahvastiku varieeruvust; latentse klassi analüüsil võivad ilmneda erinevad kliinilised tunnused, mis põhinevad näiteks põletikul.
Spetsiifiline ravi mõju põletikule ja sellele reageerimisele
Kõigil sageli määratud depressioonravi tuleb hinnata igakülgselt nende spetsiifiliste bioloogiliste mõjude alusel, mis ka arvestavad ravi katsete tõhusust. See võib võimaldada biomarkeritega ja sümptomite esitlustega seotud konstruktsioone, et prognoosida mitmesuguste antidepressantidega seotud toimetulemusi isiklikumalt ja võimalikult nii unipolaarse kui ka bipolaarse depressiooni kontekstis. See on tõenäoliselt kasulik nii uute võimalike ravimeetodite kui ka praegu näidustatud ravi korral.
Ravivastuse eeldatav kindlaksmääramine
Eespool nimetatud meetodite kasutamine võib tõenäoliselt parandada suutlikkust prognoosida resistentset seisundit edasiulatuvalt. See võib aidata kaasa autentsete ja püsivate (nt pikaajaliste) ravivastuse mõõtudele. Patsientide heaolu (nt elukvaliteet ja igapäevane toimetulek) muude kehtivate meetmete hindamine võib anda ravivõtmise terviklikuma hindamise, mis võib seostuda biomarkeritega. Kuigi bioloogiline aktiivsus üksinda ei pruugi olla võimeline eristama ravivastust mittevastavate inimeste ravivastust, võib biomarkerite andmete samaaegne mõõtmine biomarkeritega koos psühhosotsiaalsete või demograafiliste muutujatega integreerida ebapiisava ravivastuse ennustatava mudeli väljatöötamisse. Kui reageerimise ennustamiseks (kas depressioonis oleva elanikkonna või allpopulatsiooni) jaoks on välja töötatud usaldusväärne mudel ja seda valideeritakse tagasiulatuvalt, võib translatsioonisüsteem kujundada selle kohaldatavuse suure kontrollitud uuringus.
Stratifitseeritud ravi suunas
Praegu ei suunata depressiooniga patsiente süstemaatiliselt optimeeritud sekkumisprogrammi saama. Valideerimise korral võib rakendada stratifitseeritud uuringu kavandit, et ennustada reageerimatust ja / või teha kindlaks, kus astmelise hoolduse mudelis tuleb patsienti läbi viia. See võib olla kasulik nii standardiseeritud kui ka naturalistlikes raviskeemides, erinevat tüüpi sekkumiste korral. Lõppkokkuvõttes võiks välja töötada kliiniliselt elujõulise mudeli, et pakkuda inimestele kõige sobivamat ravi, et ära tunda neid, kellel tõenäoliselt areneb ravile allumatu depressioon, ning pakkuda neile patsientidele paremat hooldust ja jälgimist. Patsientidele, kellel on riskiresistentsuse risk, võib määrata samaaegse psühholoogilise ja farmakoloogilise ravi või kombineeritud farmakoteraapia. Spekulatiivse näitena võib osalejatele, kellel ei ole põletikuvastaseid tsütokiinitasemeid, osutada psühholoogilist, mitte farmakoloogilist ravi, samas kui eriti kõrge põletikuga patsientide alamrühm võib saada põletikuvastast ainet tavapärase ravi täienduseks. Sarnaselt kihistumisele võivad tulevikus olla võimalikud isikupärastatud ravivaliku strateegiad. Näiteks võib konkreetsel depressioonis isikul olla märkimisväärselt kõrge TNF? tasemed, kuid muid bioloogilisi kõrvalekaldeid pole ja kas lühiajalisest ravist TNF-iga võiks kasu olla? Antagonist.54 Isikupärastatud ravi võib hõlmata ka biomarkeri ekspressiooni jälgimist ravi ajal, et teavitada võimalikke sekkumise muutusi, vajaliku jätkuravi pikkust või varajaste ägenemiste markerite tuvastamist.
Uudsed ravi eesmärgid
Depressiooni korral võib olla tohutu hulk potentsiaalseid ravimeetodeid, mida pole piisavalt uuritud, sealhulgas uudsed või ümberkorraldatud sekkumised teistelt meditsiinivaldkondadelt. Mõned kõige populaarsemad sihtmärgid on olnud põletikuvastased ravimid, näiteks tselekoksiib (ja muud tsüklooksügenaas-2 inhibiitorid), TNF? antagonistid etanertsept ja infliksimab, minotsükliin või aspiriin. Need näivad paljutõotavad. 178 depressiooni suhtes on uuritud glükokortikoidivastaseid ühendeid, sealhulgas ketokonasool179 ja metürapooni, 180 aga mõlema kõrvalmõjude profiilil on puudusi ja metürapooni kliiniline potentsiaal on ebakindel. Mifepristoon181 ning kortikosteroidid fludrokortisoon ja spironolaktoon, 182 ning deksametasoon ja hüdrokortisoon183 võivad olla efektiivsed ka depressiooni lühiajalises ravis. Glutamaat-N-metüül-d-aspartaadi retseptori antagonistide, sealhulgas ketamiini, sihtimine võib olla depressiooni korral efektiivne ravi. 184 Omega-3 polüküllastumata rasvhapped mõjutavad põletikulist ja metaboolset aktiivsust ning näivad olevat depressiooni suhtes efektiivsed. on asjakohaste neurobioloogiliste radade kaudu antidepressantidega185
Sel viisil on antidepressantide biokeemilisi toimeid (vt jaotist Ravimid) kasutatud kliiniliste eeliste saamiseks teistes teadusharudes: eriti gastroenteroloogilised, neuroloogilised ja mittespetsiifilised sümptomihaigused. 188 Antidepressantide põletikuvastane toime võib olla osa mehhanismist need eelised. Liitiumit on soovitatud ka põletiku vähendamiseks kriitiliselt glükogeeni süntaasi kinaas-3 radade kaudu.189 Neile mõjudele keskendumine võib osutuda depressiooni biomarkerite allkirja jaoks informatiivseks ja omakorda võivad biomarkerid kujutada uue ravimi väljatöötamise asendusmarkereid.
Dr Alex Jimenez Insight
Depressioon on vaimse tervise häire, mida iseloomustavad tõsised meeleolu mõjutavad sümptomid, sealhulgas tegevuste huvi kaotamine. Hiljutised uuringud on aga leidnud, et depressiooni diagnoosimiseks võib olla võimalik rohkem kui lihtsalt patsiendi käitumishäireid. Teadlaste sõnul on patsiendi üldise tervise ja heaolu parandamiseks ülioluline, et tuvastada kergesti kättesaadavad biomarkerid, mis võivad depressiooni täpsemalt diagnoosida. Näiteks viitavad kliinilised leiud, et suure depressiivse häirega või MDD-ga inimestel on oma veres molekuli atsetüül-L-karnitiini või LAC-i madalam tase kui tervetel kontrollidel. Lõppkokkuvõttes võib depressiooni biomarkerite loomine aidata paremini määrata, kellel on haiguse tekkimise oht, ning aidata tervishoiutöötajatel määrata depressiooniga patsiendile parima ravivõimaluse.
Järeldus
Kirjandusest ilmneb, et ligikaudu kaks kolmandikku depressiooni põdevate patsientidest ei saavuta esmase ravile remissiooni ja vastuseta tõenäosus suureneb katsetatud ravimeetodite arvuga. Efektiivsete ravimeetodite pakkumisel on olulised tagajärjed individuaalsetele ja ühiskondlikele kuludele, sealhulgas pidev viletsus ja kehaline heaolu, enesetappude oht, tootlikkuse langus ja raisatud tervishoiuvarud. Depressiooni tohutu kirjandus näitab tohutut hulka biomarkereid, kellel on võimalik parandada depressiooniga inimeste ravi. Lisaks neurotransmitterite ja neuroendokriinsete markeritele, mida on paljude aastakümnete jooksul laialdaselt uuritud, on hiljutised teadmised esile tõsta põletikulist vastust (ja immuunsüsteemi üldisemalt), metaboolsetest ja kasvufaktoritest, mis on oluliselt seotud depressiooniga. Kuid ülemäära vastandlikud tõendid näitavad, et enne biomarkerite uurimistööde tegemist on depressiooniga inimeste juhtimise ja hoolduse parandamiseks mitmeid probleeme, millega tuleb tegeleda. Bioloogiliste süsteemide ülisuure keerukuse tõttu on suurte proovide ulatusliku hulga markerite samaaegne läbivaatamine märkimisväärne kasu bioloogiliste ja psühholoogiliste seisundite vastastikuse mõju avastamisel üksikisikute seas. Nii neurobioloogiliste parameetrite kui ka depressiooni kliiniliste mõõtmiste mõõtmise optimeerimine aitab tõenäoliselt paremat arusaamist. Selles ülevaates rõhutatakse ka potentsiaalselt muutvate tegurite (nagu haigus, vanus, tunnetus ja ravimid) uurimise tähtsust, et selgitada välja depressiooni bioloogia ja ravitakistuse mehhanismide ühtset arusaamist. On tõenäoline, et mõned markerid näitavad enim lubadust ennustada ravivastust või resistentsust spetsiifiliste ravimeetodite suhtes patsientide alarühmas, ning bioloogiliste ja psühholoogiliste andmete samaaegne mõõtmine võib suurendada võime neid potentsiaalsete tulemuste tuvastamiseks perspektiivselt tuvastada. Biomarkeri paneeli loomine mõjutab diagnostilise täpsuse ja prognoosi suurendamist, samuti depressiivse haiguse võimalikult varajases staadiumis ravi individuaalseid väljaarendamisi ja tõhusate uute ravivõtete väljatöötamist. Need mõjud võivad piirduda depressiooniga patsientide alarühmadega. Nende võimaluste suundumused täiendavad hiljutisi uurimisstrateegiaid kliiniliste sündroomide seostamiseks põhiosa neurobioloogiliste substraatidega. 6 Lisaks heterogeensuse vähendamisele võib see hõlbustada füüsilise ja vaimse tervise vahelise lugupidamise võrdsuse üleminekut. On selge, et kuigi on vaja palju tööd, on asjaomaste biomarkerite ja depressiivsete häirete vahelise seose kindlakstegemine märkimisväärne mõju depressiooni koormusele üksikisiku ja ühiskonna tasandil.
Tunnustused
See aruanne esindab sõltumatut uurimistööd, mida rahastavad Lõuna-Londoni Riikliku Terviseuuringute Instituudi (NIHR) biomeditsiiniliste uuringute keskus ning Maudsley NHS Foundation Trust ja Londoni King's College. Esitatud seisukohad on autorite ja mitte tingimata NHSi, NIHRi või tervishoiuministeeriumi seisukohad.
Allmärkused
Avalikustamine. AHY on viimase 3 aasta jooksul saanud tasusid Astra Zeneca (AZ), Lundbeck, Eli Lilly, Sunovioni eest rääkimise eest; Allergani, Livanova ja Lundbecki, Sunovioni, Jansseni nõustamishinnad; ja Jansseni ja Ühendkuningriigi rahastamisagentuuride (NIHR, MRC, Wellcome Trust) teadustöö toetus. AJC sai viimastel 3-aastatel Astra Zenecale (AZ) kõnelemise eest tasu, Allergani, Livanova ja Lundbeckiga konsulteerimise tasu ning Lundbecki ja Ühendkuningriigi rahastamisasutuste (NIHR, MRC, Wellcome Trust) teadustöö toetus.
Autorid ei maini selles töös muid huvide konflikte.
KokkuvõtteksKui arvukad uuringud on leidnud sadu depressiooni biomarkereid, pole paljud neist veel kindlaks teinud oma rolli depressiivses haiguses või seda, kuidas täpselt bioloogilist teavet saaks kasutada diagnoosi, ravi ja prognoosi parandamiseks. Kuid ülaltoodud artiklis vaadeldakse olemasolevat kirjandust muude protsesside käigus kaasatud biomarkerite kohta ja võrreldakse kliinilisi tulemusi depressiooni omaga. Lisaks võivad depressiooni biomarkerite uued leiud aidata depressiooni paremini diagnoosida, et jätkata paremat ravi. Riikliku biotehnoloogiaalase teabe keskuse (NCBI) viidatud teave. Meie teabe ulatus piirdub kiropraktika, samuti selgroovigastuste ja seisunditega. Teema arutamiseks küsige palun dr Jimenezilt või võtke meiega ühendust aadressil915-850-0900.
Juhib dr Alex Jimenez
Täiendavad teemad: Seljavalu
Seljavalu on üks kõige levinumaid puudeid ja töölt puuduvaid tööpäevi kogu maailmas. Tegelikkuses on seljavalu olnud ka arstikeskuse külastuste teine kõige levinum põhjus, millest arvukus oli ainult ülemiste hingamisteede nakkustega. Ligikaudu 80 protsenti elanikkonnast kogeb teatud tüüpi seljavalu vähemalt korra kogu eluea jooksul. Lülisamba on keeruline struktuur, mis koosneb teistest pehmetest kudedest koosnevate luude, liigeste, sidemete ja lihaste vahel. Selle tagajärjel tekivad vigastused ja / või raskendavad seisundid, näiteks hernide kettad, võib lõpuks põhjustada seljavalu sümptomeid. Sageli on seljavalu kõige sagedasem spordivigastus või autoõnnetusjuhtumite põhjus, kuid mõnikord võib kõige lihtsam liikumine olla valulike tulemustega. Õnneks võivad alternatiivsed ravivõimalused, nagu kiropraktiline ravi, aidata leevendada seljavalu, kasutades selleks spinaalset reguleerimist ja manuaalseid manipulatsioone, parandades lõpuks valu hõlbustamist.
4. Fava M. Raviresistentse depressiooni diagnoos ja määratlusBioli psühhiaatria2003;53(8): 649-659[PubMed]
5. Insel T, Cuthbert B, Garvey M jt. Uurimisvaldkonna kriteeriumid (RDoC): psüühikahäirete uurimise uue klassifikatsiooniraamistiku pooleOlen J psühhiaatria2010;167(7): 748-751[PubMed]
6. Kapur S, Phillips AG, Insel TR. Miks on bioloogilise psühhiaatria kliiniliste testide väljatöötamine nii kaua aega võtnud ja mida sellega teha?Mol psühhiaatria2012;17(12): 1174-1179[PubMed]
7. Gaynes BN, Warden D, Trivedi MH, Wisniewski SR, Fava M, Rush JA. Mida STAR * D meile õpetas? Depressiooniga patsientide laiaulatusliku praktilise kliinilise uuringu tulemusedPsychiatr Serv2009;60(11): 1439-1445[PubMed]
8. Fekadu A, Rane LJ, Woodersoni SC, Markopoulou K, Poon L, Cleare AJ. Ravikindla depressiooni pikaajalise tulemuse ennustamine kolmanda taseme ravisBr J psühhiaatria2012;201(5): 369-375.[PubMed]
9. Fekadu A, Woodersoni SC, Markopoulo K, Donaldson C, Papadopoulos A, Cleare AJ. Mis juhtub raviresistentse depressiooniga patsientidega? Keskmise ja pika tähtajaga uuringute süstemaatiline ülevaadeJ mõjutada ebakõla2009;116(1): 2-4[PubMed]
10. Trivedi M. Ravistrateegiad remissiooni parandamiseks ja säilitamiseks raske depressiivse häire korralDialoogid Clin Neurosci2008;10(4): 377[PMC tasuta artikkel][PubMed]
11. Fekadu A, Woodersoni SC, Markopoulou K, Cleare AJ. Raviresistentse depressiooni Maudsley etapiviis: pikemaajalise tulemuse ja sümptomite püsimise ennustamineJ Clin psühhiaatria2009;70(7): 952-957[PubMed]
12. Bennabi D, Aouizerate B, El-Hage W jt. Ravitakistuse riskifaktorid unipolaarse depressiooni korral: süstemaatiline ülevaadeJ mõjutada ebakõla2015;171: 137 141. [PubMed]
13. Serretti A, Olgiati P, Liebman MN jt. Antidepressantide reageerimise kliiniline ennustamine meeleoluhäirete korral: lineaarsed mitmemõõtmelised ja närvivõrgu mudelidPsühhiaatria res2007;152(2, 3): 223, 231.[PubMed]
14. Driessen E, Holloni SD. Meeleoluhäirete kognitiivne käitumisteraapia: efektiivsus, moderaatorid ja vahendajadPsühhiaatriakliinik Põhja-Am2010;33(3): 537-555[PMC tasuta artikkel][PubMed]
15. Cleare A, Pariante C, Young A jt. Konsensuskoosoleku liikmed Tõenduspõhised juhised depressioonihäirete raviks antidepressantidega: 2008. aasta Briti psühhofarmakoloogia suuniste ülevaade.J Psychopharmacol2015;29(5): 459-525[PubMed]
16. Tunnard C, Rane LJ, Wooderson SC jt. Lapseea raskuste mõju suitsiidile ja raviresistentse depressiooni kliinilisele kulgemiseleJ mõjutada ebakõla2014;152: 122 130. [PubMed]
17. Nemeroff CB, Heim CM, Thase ME jt. Diferentsiaalne reageerimine psühhoteraapiale võrreldes farmakoteraapiaga kroonilise raske depressiooni ja lapsepõlvetraumaga patsientidelProc Natl Acad Sci USA A.2003;100(24): 14293-14296[PMC tasuta artikkel][PubMed]
19. Tule mulle. Biomarkerite kasutamine ravivastuse prognoosimiseks raske depressiooni korral: tõendid varasematest ja praegustest uuringutestDialoogid Clin Neurosci2014;16(4): 539-544[PMC tasuta artikkel][PubMed]
20. Jani BD, McLean G, Nicholl BI jt. Depressioonisümptomitega patsientide riski hindamine ja tulemuste prognoosimine: perifeerse vere biomarkerite võimaliku rolli ülevaade.Esiosa Hum Neurosci2015;9: 18. [PMC tasuta artikkel][PubMed]
21. Suravajhala P, Kogelman LJ, Kadarmideen HN. Multimeetiline andmete integreerimine ja analüüs, kasutades süsteemigeenika lähenemisviise: meetodid ja rakendused loomakasvatuses, tervishoius ja heaolusGenet Sel Evol2016;48(1): 1[PMC tasuta artikkel][PubMed]
22. Menke A. Geeniekspressioon: antidepressantravi biomarker?Rahvusvaheline psühhiaatria2013;25(5): 579-591[PubMed]
23. Peng B, Li H, Peng XX. Funktsionaalne metaboloomika: alates biomarkerite avastamisest kuni metaboloomide ümberprogrammeerimiseniValgurakk2015;6(9): 628-637[PMC tasuta artikkel][PubMed]
24. Aagaard K, Petrosino J, Keitel W jt. Inimese mikrobiomi projekti strateegia inimese mikrobiomi terviklikuks proovivõtuks ja miks see oluline onFASEB J. 2013;27(3): 1012-1022.[PMC tasuta artikkel][PubMed]
25. Sonner Z, Wilder E, Heikenfeld J jt. Ekriinse higinäärme mikrofluidika, sealhulgas biomarkerite jaotamine, transport ja biosensatsioon.Biomikrofluidika2015;9(3): 031301.[PMC tasuta artikkel][PubMed]
26. Schmidt HD, Shelton RC, Duman RS. Depressiooni funktsionaalsed biomarkerid: diagnoosimine, ravi ja patofüsioloogiaNeuropsychopharm2011;36(12): 2375-2394[PMC tasuta artikkel][PubMed]
27. J Brand S, Moller M, H Harvey B. Ülevaade meeleolu ja psühhootiliste häirete biomarkeritest: kliiniliste ja prekliiniliste korrelatsioonide lahkamine.Curr Neuropharmacol2015;13(3): 324-368.[PMC tasuta artikkel][PubMed]
29. Fu CH, Steiner H, Costafreda SG. Depressiooni kliinilise ravivastuse ennustavad närvibiomarkerid: farmakoloogiliste ja psühholoogiliste ravimeetodite funktsionaalsete ja struktuuriliste neurokujutiste uuringute metaanalüüs.Neurobiol Dis2013;52: 75 83. [PubMed]
30. Mamdani F, Berlim M, Beaulieu M, Labbe A, Merette C, Turecki G. Geeniekspressiooni biomarkerid vastuseks tsitalopraamravi korral depressiooni korral.Tõlgi psühhiaatria2011;1(6): e13.[PMC tasuta artikkel][PubMed]
31. Smith RS. Depressiooni makrofaagide teooriaMed hüpoteesid1991;35(4): 298-306[PubMed]
32. Irwin MR, Miller AH. Depressiivsed häired ja immuunsus: 20 aastat arengut ja avastamistBrain Behav Immun2007;21(4): 374-383[PubMed]
33. Maes M, Leonard B, Myint A, Kubera M, Verkerk R. Depressiooni uus 5-HT hüpotees: rakkude vahendatud immuunaktivatsioon indutseerib indoleamiin-2,3-dioksügenaasi, mis viib madalama plasma trüptofaani ja suurenenud kahjulikud trüptofaankataboliidid (TRYCAT), mis mõlemad aitavad kaasa depressiooni tekkele.Prog Neuropsychopharmacol Biol psühhiaatria2011;35(3): 702-721.[PubMed]
34. Miller AH, Maletic V, Raison CL. Põletik ja selle rahulolematus: tsütokiinide roll raske depressiooni patofüsioloogiasBioli psühhiaatria2009;65(9): 732-741[PMC tasuta artikkel][PubMed]
35. Miller AH, Raison CL. Põletiku roll depressioonis: evolutsioonilisest imperatiivist tänapäevase ravieesmärginiNat Rev Immun2016;16(1): 22-34[PMC tasuta artikkel][PubMed]
36. Raison CL, Capuron L, Miller AH. Tsütokiinid laulavad bluusi: põletik ja depressiooni patogeneesTrends Immun2006;27(1): 24-31[PMC tasuta artikkel][PubMed]
37. Raison CL, Felger JC, Miller AH. Põletik ja raviresistentsus suure depressiooni korral: täiuslik tormPsychiatr Times2013;30(9)
38. Dowlati Y, Herrmann N, Swardfager W jt. Tsütokiinide metaanalüüs raske depressiooni korralBioli psühhiaatria2010;67(5): 446-457[PubMed]
39. Eyre HA, Air T, Pradhan A jt. Kemokiinide metaanalüüs raske depressiooni korralProg Neuropsychopharmacol Biol psühhiaatria2016;68: 1 8. [PMC tasuta artikkel][PubMed]
40. Haapakoski R, Mathieu J, Ebmeier KP, Alenius H, Kivim ki M. Interleukiinide 6 ja 1? Kumulatiivne metaanalüüs, kasvaja nekroosifaktor? ja C-reaktiivne valk raske depressiivse häirega patsientidelBrain Behav Immun2015;49: 206 215. [PMC tasuta artikkel][PubMed]
41. Howren MB, Lamkin DM, Suls J. Depressiooni seosed C-reaktiivse valgu, IL-1 ja IL-6-ga: metaanalüüs.Psühhosoom Med2009;71(2): 171-186[PubMed]
42. Liu Y, Ho RC-M, Mak A. Interleukiin (IL) -6, tuumori nekroosifaktori alfa (TNF-a) ja lahustuva interleukiin-2 retseptorid (sIL-2R) on kõrgendatud depressiivse häirega patsientidel: analüüs ja metaregressioonJ mõjutada ebakõla2012;139(3): 230-239[PubMed]
43. Strawbridge R, Arnone D, Danese A, Papadopoulos A, Herane Vives A, Cleare AJ. Põletik ja kliiniline reaktsioon depressiooni ravile: metaanalüüsEur Neuropsychopharmacol2015;25(10): 1532-1543[PubMed]
44. Farooq RK, Asghar K, Kanwal S, Zulqernain A. Põletikuliste tsütokiinide roll depressioonis: keskenduge interleukiin-1-le? (Ülevaade)Biomed Rep2017;6(1): 15-20[PMC tasuta artikkel][PubMed]
45. Cattaneo A, Ferrari C, Uher R jt. Makrofaagide migratsiooni inhibeeriva faktori ja interleukiin-1-? mRNA tase ennustab depressiooniga patsientide ravivastust täpseltInt J Neuropsychopharmacol2016;19(10): pyw045. [PMC tasuta artikkel][PubMed]
46. Baune B, Smith E, Reppermund S jt. Põletikulised biomarkerid ennustavad vananemise ajal depressiooni, kuid mitte ärevuse sümptomeid: tulevane Sydney mälu ja vananemise uuring.Psühhoneuroendokrinool2012;37(9): 1521-1530[PubMed]
47. Fornaro M, Rocchi G, Escelsior A, Contini P, Martino M. Duloksetiini saavatel depressiooniga patsientidel võivad erinevad tsütokiinide suundumused viidata erinevale bioloogilisele taustale.J mõjutada ebakõla2013;145(3): 300-307[PubMed]
48. Hernandez ME, Mendieta D, Martinez-Fong D jt. Tsirkokiini taseme varieerumine 52-nädalase SSRI-ravikuuri ajal depressiooni korralEur Neuropsychopharmacol2008;18(12): 917-924[PubMed]
49. Hannestad J, DellaGioia N, Bloch M. Antidepressantravi mõju põletikuliste tsütokiinide tasemele seerumis: metaanalüüs.Neuropsühhofarmakoloogia2011;36(12): 2452-2459.[PMC tasuta artikkel][PubMed]
50. Hiles SA, Attia J, Baker AL. Interleukiin-6, C-reaktiivse valgu ja interleukiin-10 muutused depressiooniga inimestel pärast antidepressantravi: metaanalüüs.Brain Behav Immun; Esitatakse aadressil: 17 aastakoosolek PsychoNeuroImmunology Research Society PsychoNeuroImmunology: piiriülesed haiguste vastu võitlemise valdkonnad; 2012. p. S44
51. Harley J, Luty S, Carter J, Mulder R, Joyce P. Kõrgendatud C-reaktiivne valk depressioonis: hea pikaajalise tulemuse ennustaja antidepressantidega ja halb tulemus psühhoteraapia korral.J Psychopharmacol2010;24(4): 625-626[PubMed]
52. Uher R, Tansey KE, Dew T jt. Põletikuline biomarker estsitalopraami ja nortriptüliiniga ravitud depressiooni ravi diferentsiaalse ennustajana.Olen J psühhiaatria2014;171(2): 1278-1286.[PubMed]
53. Chang HH, Lee IH, Gean PW jt. Ravivastus ja kognitiivsed häired raske depressiooni korral: seos C-reaktiivse valgugaBrain Behav Immun2012;26(1): 90-95[PubMed]
54. Raison CL, Rutherford RE, Woolwine BJ jt. Kasvaja nekroosifaktori antagonisti infliksimabi randomiseeritud kontrollitud uuring raviresistentse depressiooni korral: algsete põletikuliste biomarkerite roll.JAMA psühhiaatria2013;70(1): 31-41[PMC tasuta artikkel][PubMed]
56. Raison CL, Miller AH. Kas depressioon on põletikuline häire?Curr psühhiaatria esindaja2011;13(6): 467-475[PMC tasuta artikkel][PubMed]
57. Simon N, McNamara K, Chow C jt. Tsütokiinide kõrvalekallete üksikasjalik uurimine raske depressiooni korralEur Neuropsychopharmacol2008;18(3): 230-233[PMC tasuta artikkel][PubMed]
58. Dahl J, Ormstad H, Aass HC jt. Erinevate tsütokiinide plasmatasemed suurenevad käimasoleva depressiooni ajal ja vähenevad pärast taastumist normaalsele tasemelePsühhoneuroendokrinool2014;45: 77 86. [PubMed]
59. Stelzhammer V, Haenisch F, Chan MK jt. Esmakordselt esinenud antidepressantide raskete depressiooniga patsientide seerumiproteoomilised muutusedInt J Neuropsychopharmacol2014;17(10): 1599-1608[PubMed]
60. Liu Y, HO RCM, Mak A. Interleukiini (IL) -17 roll reumatoidartriidiga patsientide ärevuses ja depressioonis.Int J Rheum Dis. 2012;15(2): 183-187[PubMed]
61. Diniz BS, Sibille E, Ding Y jt. Hilise eluea depressiooni korral püsiva kognitiivse kahjustusega seotud plasma biosignatuur ja aju patoloogiaMol psühhiaatria2015;20(5): 594-601[PMC tasuta artikkel][PubMed]
62. Janelidze S, Ventorp F, Erhardt S jt. Muutunud kemokiini tase enesetapukatse tserebrospinaalvedelikus ja plasmasPsühhoneuroendokrinool2013;38(6): 853-862[PubMed]
63. Powell TR, Schalkwyk LC, Heffernan AL jt. Kasvajanekroosifaktor ja selle sihtmärgid põletikulises tsütokiinirajal on estsitalopraamivastuse eeldatavad transkriptoomilised biomarkerid.Eur Neuropsychopharmacol2013;23(9): 1105-1114[PubMed]
64. Wong M, Dong C, Maestre-Mesa J, Licinio J. Põletikuga seotud geenide polümorfismid on seotud vastuvõtlikkusega raskele depressioonile ja antidepressantide reaktsioonile.Mol psühhiaatria2008;13(8): 800-812[PMC tasuta artikkel][PubMed]
65. Kling MA, Alesci S, Csako G jt. Püsiv madala astme põletikueelne seisund ravimata, remissiooniga naistel, kellel on depressiivne häire, mida tõendab akuutse faasi valkude C-reaktiivse valgu ja seerumi amüloid A.Bioli psühhiaatria2007;62(4): 309-313[PMC tasuta artikkel][PubMed]
66. Schäfer M, Sarkar S, Schwarz M, Friebe A. Lahustuv rakusisese adhesiooni molekul-1 unipolaarse või bipolaarse afektiivse häirega patsientidel: tulemused pilootkatsest.Neuropsühhobiol2016;74(1): 8-14.[PubMed]
67. Dimopoulos N, Piperi C, Salonicioti A jt. Haardumolekulide plasmakontsentratsiooni tõus hilise eluea depressioonisInt J Geriatr psühhiaatria2006;21(10): 965-971[PubMed]
68. Bocchio-Chiavetto L, Bagnardi V, Zanardini R jt. Seerumi ja plasma BDNF tasemed raske depressiooni korral: replikatsiooniuuring ja metaanalüüsidMaailm J Bioli psühhiaatria2010;11(6): 763-773[PubMed]
69. Brunoni AR, Lopes M, Fregni F. Suure depressiooni ja BDNF-i taseme kliiniliste uuringute süstemaatiline ülevaade ja metaanalüüs: tagajärjed neuroplastilisuse rollile depressioonis.Int J Neuropsychopharmacol2008;11(8): 1169-1180[PubMed]
70. Molendijk M, Spinhoven P, Polak M, Bus B, Penninx B, Elzinga B. Seerumi BDNF-i kontsentratsioonid depressiooni perifeersete ilmingutena: tõendid süstemaatilisest ülevaatest ja metaanalüüsidest 179 assotsiatsiooni kohta.Mol psühhiaatria2014;19(7): 791-800[PubMed]
71. Sen S, Duman R, Sanacora G. Seerumi ajupõhine neurotroofne faktor, depressioon ja antidepressandid: metaanalüüsid ja tagajärjed.Bioli psühhiaatria2008;64(6): 527-532[PMC tasuta artikkel][PubMed]
72. Zhou L, Xiong J, Lim Y jt. Vere proBDNF ja selle retseptorite reguleerimine raske depressiooni korralJ mõjutada ebakõla2013;150(3): 776-784[PubMed]
73. Chen YW, Lin PY, Tu KY, Cheng YS, Wu CK, Tseng PT. Märkimisväärselt madalam närvikasvufaktori tase raske depressiivse häirega patsientidel kui tervetel isikutel: metaanalüüs ja süsteemne ülevaade.Neuropsühhiaatriline haigus2014;11: 925 933. [PMC tasuta artikkel][PubMed]
74. Lin PY, Tseng PT. Depressiooniga patsientide gliaalrakuliinist tuletatud neurotroofse faktori taseme langus: metaanalüütiline uuring.J Psychiatr Res2015;63: 20 27. [PubMed]
75. Warner-Schmidt JL, Duman RS. VEGF kui depressiooni terapeutilise sekkumise potentsiaalne sihtmärkCurr Op Pharmacol2008;8(1): 14-19[PMC tasuta artikkel][PubMed]
76. Carvalho AF, Khler CA, McIntyre RS jt. Perifeerse vaskulaarse endoteeli kasvufaktor kui uudne depressiooni biomarker: metaanalüüsPsühhoneuroendokrinool2015;62: 18 26. [PubMed]
78. Carvalho L, Torre J, Papadopoulos A jt. Antidepressantide kliinilise terapeutilise kasu puudumine on seotud põletikulise süsteemi üldise aktiveerimisegaJ mõjutada ebakõla2013;148(1): 136-140[PubMed]
79. Clark-Raymond A, Meresh E, Hoppensteadt D jt. Vaskulaarne endoteeli kasvufaktor: potentsiaalne ravivastuse ennustaja raske depressiooni korralMaailm J Bioli psühhiaatria2015: 1 11[PubMed]
80. Isung J, Mobarrez F, Nordstrm P, Sberg M, Jokinen J. Lõppenud enesetapuga seotud madal vereplasma veresoonte endoteeli kasvufaktor (VEGF).Maailm J Bioli psühhiaatria2012;13(6): 468-473[PubMed]
82. Szcz? Sny E,? Lusarczyk J, G? Ombik K jt. IGF-1 võimalik panus depressiivsesse häiressePharmacol Rep2013;65(6): 1622-1631[PubMed]
83. Tu KY, Wu MK, Chen YW jt. Märkimisväärselt kõrgem perifeerse insuliinilaadse kasvufaktori-1 tase depressiooni või bipolaarse häire korral tervetel kontrollgruppidel: metaanalüüs ja ülevaade PRISMA juhendi alusel.Med2016;95(4): e2411. [PMC tasuta artikkel][PubMed]
84. Wu CK, Tseng PT, Chen YW, Tu KY, Lin PY. Märkimisväärselt kõrgem perifeerse fibroblasti kasvufaktori-2 tase raske depressiivse häirega patsientidel: esialgne metaanalüüs vastavalt MOOSE juhistele.Med2016;95(33): e4563. [PMC tasuta artikkel][PubMed]
85. He S, Zhang T, Hong B jt. Seerumi fibroblastide kasvufaktori-2 taseme langus raske depressiivse häirega patsientidel enne ja pärast raviNeurosci Lett2014;579: 168 172. [PubMed]
86. Dwivedi Y, Rizavi HS, Conley RR, Roberts RC, Tamminga CA, Pandey GN. Ajust pärineva neurotroofse faktori ja retseptori türosiinikinaasi B muutunud geeniekspressioon enesetapjate subjektide surmajärgses ajus.Arch Gen psühhiaatria2003;60(8): 804-815[PubMed]
87. Srikanthan K, Feyh A, Visweshwar H, Shapiro JI, Sodhi K. Metaboolse sündroomi biomarkerite süstemaatiline ülevaade: Lääne-Virginia elanikkonna varajase avastamise, haldamise ja riskikihistumise paneel.Int J Med Sci2016;13(1): 25[PMC tasuta artikkel][PubMed]
88. Lu XY. Depressiooni leptiini hüpotees: potentsiaalne seos meeleoluhäirete ja rasvumise vahel?Curr Op Pharmacol2007;7(6): 648-652[PMC tasuta artikkel][PubMed]
89. Wittekind DA, Kluge M. Ghrelin psühhiaatriliste häirete korral. ÜlevaadePsühhoneuroendokrinool2015;52: 176 194. [PubMed]
90. Kan C, Silva N, Golden SH jt. Depressiooni ja insuliiniresistentsuse seose süstemaatiline ülevaade ja metaanalüüsDiabeedi hooldus2013;36(2): 480-489[PMC tasuta artikkel][PubMed]
91. Liu X, Li J, Zheng P jt. Plasma lipidoomika näitab võimalikke depressiivse häire lipiidimarkereidAnal Bioanal Chem2016;408(23): 6497-6507[PubMed]
92. Lustman PJ, Anderson RJ, Freedland KE, De Groot M, Carney RM, Clouse RE. Depressioon ja halb glükeemiline kontroll: kirjanduse metaanalüütiline ülevaadeDiabeedi hooldus2000;23(7): 934-942[PubMed]
93. Maes M. Tõendid immuunvastuse kohta depressiooni korral: ülevaade ja hüpoteesProg NeuroPsychopharmacol Biol psühhiaatria1995;19(1): 11-38[PubMed]
94. Zheng H, Zheng P, Zhao L jt. Suure depressiooni ennustav diagnoosimine NMR-põhise metaboloomika ja väikseimate ruutude abil vektormasinagaClinica Chimica Acta2017;464: 223-227.[PubMed]
95. Xia Q, Wang G, Wang H, Xie Z, Fang Y, Li Y. Uuring glükoosi ja lipiidide metabolismist esimese episoodi depressiooniga patsientidel.J Clin psühhiaatria2009;19: 241-243.
96. Kaufman J, DeLorenzo C, Choudhury S, Parsey RV. 5-HT 1A retseptor suure depressiivse häire korralEur Neuropsühhofarmakoloogia2016;26(3): 397-410[PMC tasuta artikkel][PubMed]
98. Salamone JD, Correa M, Yohn S, Cruz LL, San Miguel N, Alatorre L. Pingutusega seotud valikukäitumise farmakoloogia: dopamiin, depressioon ja individuaalsed erinevused.Käitumisprotsessid2016;127: 3 17. [PubMed]
100. Popa D, Cerdan J, Reprant C jt. 5-HT väljavoolu pikisuunaline uuring kroonilise fluoksetiinravi ajal, kasutades uut kroonilise mikrodialüüsi tehnikat väga emotsionaalses hiire tüves.Eur J Pharmacol2010;628(1): 83-90[PubMed]
101. Atake K, Yoshimura R, Hori H jt. Duloksetiin, selektiivne noradrenaliini tagasihaarde inhibiitor, suurendas raske depressiivse häirega patsientidel 3-metoksü-4-hüdroksüfenüülglükooli, kuid mitte homovanilliinhappe taset plasmas.Clin Psychopharmacol Neurosci2014;12(1): 37-40[PMC tasuta artikkel][PubMed]
102. Ueda N, Yoshimura R, Shinkai K, Nakamura J. Katehhoolamiini metaboliitide plasmatase ennustab reaktsiooni sulpiriidile või fluvoksamiinile depressiooni korral.Farmakopsühhiaatria2002;35(05): 175-181.[PubMed]
103. Yamana M, Atake K, Katsuki A, Hori H, Yoshimura R. Vere bioloogilised markerid estsitalopraamivastuse prognoosimiseks raske depressiivse häirega patsientidel: eeluuring.J Depresseerige ärevust2016;5: 222.
104. Parker KJ, Schatzbergi AF, Lyoni DM. Hüperkortisolismi neuroendokriinsed aspektid raske depressiooni korralHorm käitumine2003;43(1): 60-66[PubMed]
105. Stetler C, Miller GE. Depressioon ja hüpotalamuse-hüpofüüsi-neerupealiste aktiveerimine: kvantitatiivne kokkuvõte nelja aastakümne uuringustPsühhosoom Med2011;73(2): 114-126[PubMed]
106. Herane Vives A, De Angel V, Papadopoulos A jt. Kortisooli, stressi ja psühhiaatriliste haiguste seos: uued teadmised juuste analüüsi abilJ Psychiatr Res2015;70: 38 49. [PubMed]
108. Anacker C, Zunszain PA, Carvalho LA, Pariante CM. Glükokortikoidiretseptor: depressiooni ja antidepressantide ravi pöördepunkt?Psühhoneuroendokrinoloogia2011;36(3): 415-425[PMC tasuta artikkel][PubMed]
109. Markopoulou K, Papadopoulos A, Juruena MF, Poon L, Pariante CM, Cleare AJ. Kortisooli / DHEA suhe raviresistentses depressioonisPsühhoneuroendokrinool2009;34(1): 19-26[PubMed]
110. Joffe RT, Pearce EN, Hennessey JV, Ryan JJ, Stern RA. Subkliiniline hüpotüreoidism, meeleolu ja tunnetus vanematel täiskasvanutel: ülevaadeInt J Geriatr psühhiaatria2013;28(2): 111-118[PMC tasuta artikkel][PubMed]
111. Duval F, Mokrani MC, Erb A jt. Kronobioloogiline hüpotalamuse hüpofüüsi kilpnäärme telje seisund ja antidepressandi tulemus raske depressiooni korralPsühhoneuroendokrinool2015;59: 71 80. [PubMed]
112. Marsden W. Sünaptiline plastilisus depressioonis: molekulaarne, rakuline ja funktsionaalne korrelatsioonProg Neuropsychopharmacol Biol psühhiaatria2013;43: 168 184. [PubMed]
113. Duman RS, Voleti B. Depressiooni patofüsioloogia ja ravi aluseks olevad signaalirajad: uudsed kiiretoimeliste ainete mehhanismid.Trendid Neurosci2012;35(1): 47-56.[PMC tasuta artikkel][PubMed]
114. Ripke S, Wray NR, Lewis CM jt. Suure depressiivse häire kogu genoomi hõlmavate uuringute megaanalüüsMol psühhiaatria2013;18(4): 497-511[PMC tasuta artikkel][PubMed]
115. Mullins N, Power R, Fisher H jt. Polügeensed koostoimed keskkonna raskustega raske depressiivse häire etioloogiasPsychol Med2016;46(04): 759-770[PMC tasuta artikkel][PubMed]
116. Lewis S. Neuroloogilised häired: telomeerid ja depressioonNat Rev Neurosci2014;15(10): 632.[PubMed]
117. Lindqvist D, Epel ES, Mellon SH jt. Psühhiaatrilised häired ja leukotsüütide telomeeride pikkus: vaimse haiguse ja raku vananemisega seotud mehhanismidNeurosci Biobehav Rev.2015;55: 333 364. [PMC tasuta artikkel][PubMed]
118. McCall WV. Puhketegevuse biomarker, et ennustada reaktsiooni SSRI-dele depressiooni korralJ Psychiatr Res2015;64: 19 22. [PMC tasuta artikkel][PubMed]
120. Foster JA, Neufeld K-AM. Soole aju telg: kuidas mikrobioom mõjutab ärevust ja depressiooniTrendid Neurosci2013;36(5): 305-312[PubMed]
121. Quattrocki E, Baird A, Yurgelun-Todd D. Suitsetamise ja depressiooni vahelise seose bioloogilised aspektid.Harv Rev psühhiaatria2000;8(3): 99-110[PubMed]
122. Maes M, Kubera M, Obuchowiczwa E, Goehler L, Brzeszcz J. Depressiooni mitu kaasuvat haigust, mida seletatakse (neuro) põletikuliste ning oksüdatiivsete ja nitrosatiivsete stressiteedega.Neuro Endocrinol Lett2011;32(1): 7-24[PubMed]
123. Miller G, Rohleder N, Cole SW. Krooniline inimestevaheline stress ennustab pro- ja põletikuvastaste signaaliradade aktiveerimist kuus kuud hiljemPsühhosoom Med2009;71(1): 57[PMC tasuta artikkel][PubMed]
124. Steptoe A, Hamer M, Chida Y. Ägeda psühholoogilise stressi mõju ringlevatele põletikulistele teguritele inimestel: ülevaade ja metaanalüüs.Brain Behav Immun2007;21(7): 901-912[PubMed]
125. Danese A, Moffitt TE, Harrington H jt. Lapsepõlve ebasoodsad kogemused ja täiskasvanute vanusega seotud haiguste riskifaktorid: depressioon, põletik ja metaboolsete riskimarkerite klastrid.Arch Pediatr Adolesc Med2009;163(12): 1135-1143[PMC tasuta artikkel][PubMed]
126. Danese A, Pariante CM, Caspi A, Taylor A, Poulton R. Lapsepõlves väärkohtlemine ennustab elukursuse uuringus täiskasvanute põletikku.Proc Natl Acad Sci USA A.2007;104(4): 1319-1324[PMC tasuta artikkel][PubMed]
127. Danese A, Caspi A, Williams B jt. Stressi bioloogiline kinnistamine lapsepõlves põletikuliste protsesside kauduMol psühhiaatria2011;16(3): 244-246[PMC tasuta artikkel][PubMed]
128. Suzuki A, Poon L, Kumari V, Cleare AJ. Lapsepõlvetraumale järgnenud emotsionaalse näotöötluse hirm kallutatus, mis on vastupidavuse ja depressiooni suhtes haavatavuse markerLapse halb kohtlemine2015;20(4): 240-250[PubMed]
129. Strawbridge R, noor AH. HPA telg ja kognitiivne düsregulatsioon meeleoluhäirete korral. Koos: McIntyre RS, Cha DS, toimetajadKognitiivsed häired raske depressiivse häire korral: kliiniline tähtsus, bioloogilised substraadid ja ravivõimalused.Cambridge: Cambridge University Press; 2016. lk 179 193.
130. Keller J, Gomez R, Williams G jt. HPA telg suure depressiooni korral: kortisool, kliiniline sümptomatoloogia ja geneetiline variatsioon ennustavad tunnetustMol psühhiaatria2016 16. august; pEpub[PMC tasuta artikkel][PubMed]
131. Hanson ND, Owens MJ, Nemeroff CB. Depressioon, antidepressandid ja neurogenees: kriitiline ümberhindamineNeuropsychopharmacol2011;36(13): 2589-2602[PMC tasuta artikkel][PubMed]
132. Chen Y, Baram TZ. Selle mõistmise poole, kuidas varase elu stress kognitiivseid ja emotsionaalseid ajuvõrke ümber programmeeribNeuropsychopharmacol2015;41(1): 197-206[PMC tasuta artikkel][PubMed]
133. Porter RJ, Gallagher P, Thompson JM, Young AH. Neurokognitiivsed häired ravimivabadel patsientidel, kellel on suur depressiivne häireBr J psühhiaatria2003;182: 214 220. [PubMed]
134. Gallagher P, Robinson L, Gray J, Young A, Porter R. Neurokognitiivne funktsioon pärast remissiooni peamise depressiivse häire korral: potentsiaalne ravivastuse marker?Aust NZJ psühhiaatria2007;41(1): 54-61[PubMed]
136. Bckman L, Nyberg L, Lindenberger U, Li SC, Farde L. Korrelatiivne kolmkõla vananemise, dopamiini ja tunnetuse seas: hetkeseis ja tulevikuväljavaated.Neurosci Biobehav Rev.2006;30(6): 791-807[PubMed]
137. Allison DJ, Ditor DS. Depressiooni ja kognitiivsete häirete levinud põletikuline etioloogia: terapeutiline sihtmärkJ närvipõletik2014;11: 151. [PMC tasuta artikkel][PubMed]
138. Rosenblat JD, Brietzke E, Mansur RB, Maruschak NA, Lee Y, McIntyre RS. Põletik kui kognitiivsete häirete neurobioloogiline substraat bipolaarse häire korral: tõendid, patofüsioloogia ja tagajärjed ravile.J mõjutada ebakõla2015;188: 149 159. [PubMed]
139. Krogh J, Benros ME, Jrgensen MB, Vesterager L, Elfving B, Nordentoft M. Depressioonisümptomite, kognitiivsete funktsioonide ja põletiku seos raske depressiooni korral.Brain Behav Immun2014;35: 70 76. [PubMed]
140. Soares CN, Zitek B. Reproduktiivhormooni tundlikkus ja depressiooni oht kogu naise elutsüklis: haavatavuse pidevus?J Psühhiaatria Neurosci2008;33(4): 331[PMC tasuta artikkel][PubMed]
141. Hiles SA, Baker AL, de Malmanche T, Attia J. Depressiooniga ja depressioonita inimeste IL-6 ja IL-10 erinevuste metaanalüüs: heterogeensuse põhjuste uurimine.Brain Behav Immun2012;26(7): 1180-1188[PubMed]
142. Fontana L, Eagon JC, Trujillo ME, Scherer PE, Klein S. Vistseraalse rasva adipokiini sekretsioon on rasvunud inimestel seotud süsteemse põletikuga.Diabeet2007;56(4): 1010-1013[PubMed]
143. Divani AA, Luo X, Datta YH, Flaherty JD, Panoskaltsis-Mortari A. Suukaudsete ja tupe hormonaalsete kontratseptiivide mõju põletikulistele vere biomarkeritele.Vahendajad Põletik2015;2015: 379501.[PMC tasuta artikkel][PubMed]
144. Ramsey JM, Cooper JD, Penninx BW, Bahn S. Seksi ja naise hormonaalse seisundiga seerumi biomarkerite variatsioon: mõju kliinilistele testidele.Sci Rep2016;6: 26947. [PMC tasuta artikkel][PubMed]
145. Eyre H, Lavretsky H, Kartika J, Qassim A, Baune B. Antidepressantide klasside moduleeriv toime depressiooni kaasasündinud ja adaptiivsele immuunsüsteemile.Farmakopsühhiaatria2016;49(3): 85-96.[PMC tasuta artikkel][PubMed]
146. Hiles SA, Baker AL, de Malmanche T, Attia J. Interleukiin-6, C-reaktiivne valk ja interleukiin-10 pärast antidepressantravi depressiooniga inimestel: metaanalüüs.Psychol Med2012;42(10): 2015-2026[PubMed]
148. Artigas F. antidepressantidega seotud serotoniini retseptoridPharmacol Ther2013;137(1): 119-131[PubMed]
149. Lee BH, Kim YK. BDNF-i rollid raske depressiooni patofüsioloogias ja antidepressantide ravisPsühhiaatria uurimine2010;7(4): 231-235[PMC tasuta artikkel][PubMed]
150. Hashimoto K. Põletikulised biomarkerid antidepressantide reaktsiooni diferentsiaalse ennustajanaInt J Mol Sci2015;16(4): 7796-7801[PMC tasuta artikkel][PubMed]
151. Goldberg D. Peamise depressiooni heterogeensusMaailma psühhiaatria2011;10(3): 226-228.[PMC tasuta artikkel][PubMed]
152. Arnow BA, Blasey C, Williams LM jt. Depressiooni alatüübid antidepressantide reageerimise ennustamisel: aruanne uuringust iSPOT-DOlen J psühhiaatria2015;172(8): 743-750[PubMed]
153. Kunugi H, Hori H, Ogawa S. Biokeemilised markerid, mis allutavad suurt depressiivset häiretPsühhiaatriakliinik Neurosci2015;69(10): 597-608[PubMed]
154. Baune B, Stuart M, Gilmour A jt. Depressiooni alatüüpide ja südame-veresoonkonna haiguste seos: bioloogiliste mudelite süstemaatiline ülevaadeTõlgi psühhiaatria2012;2(3): e92.[PMC tasuta artikkel][PubMed]
155. Vogelzangs N, Duivis HE, Beekman AT jt. Depressiivsete häirete, depressiooni tunnuste ja antidepressantide seos põletikugaTõlgi psühhiaatria2012;2: e79.[PMC tasuta artikkel][PubMed]
156. Lamers F, Vogelzangs N, Merikangas K, De Jonge P, Beekman A, Penninx B. Tõendid HPA-telje funktsiooni, põletiku ja metaboolse sündroomi erinevast rollist melanhoolses versus ebatüüpilises depressioonis.Mol psühhiaatria2013;18(6): 692-699[PubMed]
157. Penninx BW, Milaneschi Y, Lamers F, Vogelzangs N. Depressiooni somaatiliste tagajärgede mõistmine: bioloogilised mehhanismid ja depressiooni sümptomite profiili roll.BMC Med2013;11(1): 1.[PMC tasuta artikkel][PubMed]
158. Capuron L, Su S, Miller AH jt. Depressiivsed sümptomid ja metaboolne sündroom: kas põletik on aluseks?Bioli psühhiaatria2008;64(10): 896-900[PMC tasuta artikkel][PubMed]
159. Dantzer R, Connor JC, Freund GG, Johnson RW, Kelley KW. Põletikust haiguse ja depressioonini: kui immuunsüsteem allutab ajuNat Rev Neurosci2008;9(1): 46-56.[PMC tasuta artikkel][PubMed]
160. Maes M, Berk M, Goehler L jt. Depressioon ja haigusega seotud käitumine on Januse näo vastused jagatud põletikuradadeleBMC Med2012;10: 66. [PMC tasuta artikkel][PubMed]
161. Merikangas KR, Jin R, He JP jt. Bipolaarse spektrihäire levimus ja korrelatsioon maailma vaimse tervise uuringu algatuselArch Gen psühhiaatria2011;68(3): 241-251[PMC tasuta artikkel][PubMed]
162. Hirschfeld RM, Lewis L, Vornik LA. Bipolaarse häire tajud ja mõju: kui kaugele oleme tegelikult jõudnud? Bipolaarse häirega inimeste 2000. aasta riikliku depressiivse ja maniakaal-depressiivse ühenduse uuringu tulemusedJ Clin psühhiaatria2003;64(2): 161-174[PubMed]
163. Young AH, MacPherson H. Bipolaarse häire tuvastamineBr J psühhiaatria2011;199(1): 3-4.[PubMed]
164. Hringer PA, Perlis RH. Bipolaarse häire ja raske depressiivse häire eristaminePsühhiaatriakliinik Põhja-Am2016;39(1): 1-10[PubMed]
165. Becking K, Spijker AT, Hoencamp E, Penninx BW, Schoevers RA, Boschloo L. Hüpotalamuse-hüpofüüsi-neerupealise telje häired ja unipolaarse ja bipolaarse depressiooni episoodide eristamine.PLoS One2015;10(7): e0133898. [PMC tasuta artikkel][PubMed]
166. Huang TL, Lin FC. Kõrge tundlikkusega C-reaktiivse valgu tase raske depressiooni ja bipolaarse maaniaga patsientidelProg NeuroPsychopharmacol Biol psühhiaatria2007;31(2): 370-372[PubMed]
167. Angst J, Gamma A, Endrass J. Bipolaarse ja depressioonispektri riskiteguridActa Psychiatr Scand2003;418: 15 19. [PubMed]
168. Fekadu A, Wooderson S, Donaldson C jt. Mitmemõõtmeline tööriist depressiooni raviresistentsuse kvantifitseerimiseks: Maudsley lavastusmeetodJ Clin psühhiaatria2009;70(2): 177[PubMed]
169. Papakostas G, Shelton R, Kinrys G jt. Suurepressiivse häire mitmeanalüüsi seerumipõhise bioloogilise diagnostilise testi hindamine: piloot- ja replikatsiooniuuring.Mol psühhiaatria2013;18(3): 332-339[PubMed]
170. Fan J, Han F, Liu H. Suurandmete analüüsi väljakutsedNatl Sci Rev.2014;1(2): 293-314.[PMC tasuta artikkel][PubMed]
171. Li L, Jiang H, Qiu Y, Ching WK, Vassiliadis VS. Metaboliitide biomarkerite avastamine: vooanalüüs ja reaktsioon-reaktsioonivõrgu lähenemineBMC Syst Biol2013;7(Varustus 2): S13[PMC tasuta artikkel][PubMed]
172. Patel MJ, Khalaf A, Aizenstein HJ. Depressiooni uurimine pildistamise ja masinõppe meetodite abilNeuroImage Clin2016;10: 115 123. [PMC tasuta artikkel][PubMed]
173. Lanquillon S, Krieg JC, Bening-Abu-Shach U, Vedder H. Tsütokiinide tootmine ja ravivastus suure depressiivse häire korral.Neuropsychopharmacol2000;22(4): 370-379[PubMed]
175. Hidalgo-Mazzei D, Murru A, Reinares M, Vieta E, Colom F. Vaimse tervise suured andmed: keeruline killustatud tulevik.Maailma psühhiaatria2016;15(2): 186-187[PMC tasuta artikkel][PubMed]
176. Konsortsium C-DGotPG Viie peamise psühhiaatrilise häire jagatud mõjudega riskilookuste kindlakstegemine: kogu genoomi hõlmav analüüs.Lancet2013;381(9875): 1371-1379[PMC tasuta artikkel][PubMed]
177. Dipnall JF, Pasco JA, Berk M jt. Andmekaevanduse, masinõppe ja traditsioonilise statistika ühendamine depressiooniga seotud biomarkerite tuvastamiseksPLoS One2016;11(2): e0148195. [PMC tasuta artikkel][PubMed]
178. Khler O, Benros ME, Nordentoft M jt. Põletikuvastase ravi mõju depressioonile, depressioonisümptomitele ja kõrvaltoimetele: randomiseeritud kliiniliste uuringute süstemaatiline ülevaade ja metaanalüüs.JAMA psühhiaatria2014;71(12): 1381-1391[PubMed]
179. Wolkowitz OM, Reus VI, Chan T jt. Depressiooni glükokortikoidivastane ravi: topeltpime ketokonasoolBioli psühhiaatria1999;45(8): 1070-1074[PubMed]
180. McAllister-Williams RH, Anderson IM, Finkelmeyer A jt. Antidepressantide suurendamine metürapooniga raviresistentse depressiooni korral (ADD uuring): topeltpime, randomiseeritud, platseebokontrolliga uuring.Lanceti psühhiaatria2016;3(2): 117-127[PubMed]
181. Gallagher P, noor AH. Mifepristooni (RU-486) depressiooni ja psühhoosi ravi: terapeutiliste tagajärgede ülevaadeNeuropsühhiaatriline haigus2006;2(1): 33-42[PMC tasuta artikkel][PubMed]
182. Otte C, Hinkelmann K, Moritz S jt. Mineralokortikoidiretseptori moduleerimine depressiooni lisaravina: randomiseeritud, topeltpime, platseebokontrolliga uuring.J Psychiatr Res2010;44(6): 339-346[PubMed]
186. Kim S, Bae K, Kim J jt. Statiinide kasutamine depressiooni raviks ägeda koronaarsündroomiga patsientidelTõlgi psühhiaatria2015;5(8): e620. [PMC tasuta artikkel][PubMed]
187. Shishehbor MH, Brennan ML, Aviles RJ jt. Statiinid soodustavad spetsiifiliste põletikuliste radade kaudu tugevat süsteemset antioksüdantset toimetTiraaž2003;108(4): 426-431[PubMed]
188. Mercier A, Auger-Aubin I, Lebeau JP jt. Tõendid antidepressantide väljakirjutamise kohta psühhiaatriliste haiguste korral esmatasandi arstiabis: juhiste ja süstemaatiliste ülevaadete analüüs.BMC perepraktika2013;14(1): 55[PMC tasuta artikkel][PubMed]
189. Freland L, Beaulieu JM. GSK3 pärssimine liitiumiga, üksikutest molekulidest signaalivõrkudeniEsiosa Mol Neurosci2012;5: 14. [PMC tasuta artikkel][PubMed]
190. Horowitz MA, Zunszain, PA. Neuroimmuunsed ja neuroendokriinsed kõrvalekalded depressioonis: sama mündi kaks külgeAnn NY Acad Sci2015;1351(1): 68-79[PubMed]
191. Juruena MF, Cleare AJ. Ebatüüpilise depressiooni, hooajalise afektiivse häire ja kroonilise väsimussündroomi kattuvusRev Bras Psiquiatr2007;29: S19 S26. [PubMed]
192. Castr n E, Kojima M. ajupõhine neurotroofne faktor meeleoluhäirete ja antidepressantravi korral.Neurobiol Dis2017;97(Pt B): 119[PubMed]
193. Pan A, Keum N, Okereke OI jt. Depressiooni ja metaboolse sündroomi kahesuunaline seos, epidemioloogiliste uuringute süstemaatiline ülevaade ja metaanalüüsDiabeedi hooldus2012;35(5): 1171-1180[PMC tasuta artikkel][PubMed]
194. Carvalho AF, Rocha DQ, McIntyre RS jt. Adipokiinid kui tekkivad depressiooni biomarkerid: süstemaatiline ülevaade ja metaanalüüsJ Psühhiaatria Res2014;59: 28 37. [PubMed]
195. Wise T, Cleare AJ, Herane A, Young AH, Arnone D. Neurokujutiste diagnostiline ja terapeutiline kasulikkus depressioonis: ülevaade.Neuropsühhiaatriline haigus2014;10: 1509-1522.[PMC tasuta artikkel][PubMed]
196. Tamatam A, Khanum F, Bawa AS. Depressiooni geneetilised biomarkeridIndiaanlane J Hum Genet2012;18(1): 20[PMC tasuta artikkel][PubMed]
197. Yoshimura R, Nakamura J, Shinkai K, Ueda N. Kliiniline reaktsioon antidepressantravi ja 3-metoksü-4-hüdroksüfenüülglükooli tasemele: mini ülevaade.Prog Neuropsychopharmacol Biol psühhiaatria2004;28(4): 611-616[PubMed]
198. Pierscionek T, Adekunte O, Watson S, Ferrier N, Alabi A. Kortikosteroidide roll antidepressantide ravis.ChronoPhys Ther2014;4: 87-98.
199. Hage MP, Azar ST. Seos kilpnäärme funktsiooni ja depressiooni vahelJ Kilpnäärme res2012;2012: 590648. [PMC tasuta artikkel][PubMed]
200. Dunn EC, Brown RC, Dai Y jt. Depressiooni geneetilised määrajad: hiljutised leiud ja edasised suunadHarv Rev psühhiaatria2015;23(1): 1[PMC tasuta artikkel][PubMed]
201. Yang CC, Hsu YL. Füüsilise aktiivsuse jälgimiseks mõeldud kiirendusmõõtmel põhinevate kantavate liikumisandurite ülevaadeAndurid2010;10(8): 7772-7788[PMC tasuta artikkel][PubMed]
Artriit valu on keeruline nähtus, mis hõlmab keerulist neurofüsioloogilist töötlemist valuraja kõigil tasanditel. Liigesevalu leevendamiseks saadaolevad ravivõimalused on üsna piiratud ja enamik artriidiga patsiente teatab praeguste ravimeetoditega ainult mõõdukast valu leevendust. Lihas-skeleti valu tekitavate neuraalsete mehhanismide parem mõistmine ja uute sihtmärkide tuvastamine aitab välja töötada tulevasi farmakoloogilisi ravimeetodeid. Selles artiklis antakse ülevaade viimastest uuringutest liigesevalu soodustavate tegurite kohta ning käsitletakse selliseid valdkondi nagu kannabinoidid, proteinaasiga aktiveeritud retseptorid, naatriumikanalid, tsütokiinid ja transientsete retseptorite potentsiaalikanalid. Samuti arutatakse esilekerkivat hüpoteesi, et osteoartriidil võib olla neuropaatiline komponent.
Sissejuhatus
Maailma tervishoiuorganisatsioon liigitab luu-ja lihaskonna vaevused kõige sagedasemaks puuete põhjuseks tänapäeva maailmas, mõjutades üks kolmest täiskasvanust [1]. Veelgi murettekitavam on see, et nende haiguste esinemissagedus tõuseb, samal ajal kui meie teadmised nende põhjuste kohta on üsna algelised.
Joonis 1 Skeem, mis illustreerib mõningaid sihtmärke, mis teadaolevalt moduleerivad liigesevalu. Neuromodulaatoreid saab vabastada nii närvilõpmetest kui ka nuumrakkudest ja makrofaagidest, et muuta aferentset mehaanilist tundlikkust. Endovanilloidid, hape ja kahjulik kuumus võivad aktiveerida retseptori potentsiaali mööduvaid vanilloidi tüüpi 1 (TRPV1) ioonikanaleid, mille tulemuseks on algogeense aine P (SP) vabanemine, mis seejärel seondub neurokiniin-1 (NK1) retseptoritega. Proteaasid võivad lõhustada ja stimuleerida proteaasiga aktiveeritud retseptoreid (PAR). Siiani on näidatud, et PAR2 ja PAR4 sensibiliseerivad liigese primaarseid aferente. Endokannabinoid-anandamiid (AE) toodetakse nõudmisel ja sünteesitakse N-arahhidonoüülfosfatidüületanoolamiinist (NAPE) fosfolipaaside ensümaatilise toimega. Seejärel seondub osa AE-st kannabinoid-1 (CB1) retseptoritega, mis viib neuronite desensibiliseerumiseni. Seondumata AE imendub kiiresti anandamiidmembraani transporterisse (AMT), enne kui see laguneb rasvhappeamiidi hüdrolaasi (FAAH) toimel etanoolamiiniks (Et) ja arahhidoonhappeks (AA). Tsütokiinid tuumori nekroosifaktor-a (TNF-a), interleukiin-6 (IL-6) ja interleukiin-1-beeta (IL-1a) võivad seostuda oma vastavate retseptoritega, et tugevdada valu edasikandumist. Lõpuks osalevad neuronite sensibiliseerimises tetrodotoksiini (TTX) resistentsed naatriumikanalid (Nav1.8).
Patsiendid igatsevad nende eest kroonilise valu kaduda; praegu välja kirjutatud valuvaigistid on aga suures osas ebaefektiivsed ja nendega kaasneb suur hulk soovimatuid kõrvalmõjusid. Seetõttu kannatavad miljonid inimesed kogu maailmas liigesevalu kurnavate tagajärgede all, millele ei ole rahuldavat ravi olemas [2].
Rohkem kui 100 erineva artriidi vormi puhul on kõige levinum osteoartriit (OA). OA on progresseeruv degeneratiivne liigesehaigus, mis põhjustab kroonilist valu ja funktsiooni kaotust. Tavaliselt on OA liigese võimetus kahjustusi tõhusalt parandada vastuseks sellele, et sellele rakendatakse liigseid jõude. Bioloogilised ja psühhosotsiaalsed tegurid, mis hõlmavad kroonilist OA valu, ei ole hästi teada, kuigi käimasolevad uuringud selgitavad lahti haiguse sümptomite keerukuse [2]. Praegused ravimid, nagu mittesteroidsed põletikuvastased ravimid (MSPVA-d), pakuvad mõningast sümptomaatilist leevendust, vähendades valu lühikese aja jooksul, kuid ei leevenda valu kogu patsiendi eluea jooksul. Lisaks ei saa suurtes annustes MSPVA-sid võtta korduvalt paljude aastate jooksul, kuna see võib põhjustada neerutoksilisust ja seedetrakti verejooksu.
Traditsiooniliselt on artriidi uuringud keskendunud suures osas liigesekõhrele kui peamisele sihtmärgile uudsete OA-ravimite väljatöötamisel haiguse modifitseerimiseks. See kondrogeenne fookus on heitnud uut valgust keerukatele biokeemilistele ja biomehaanilistele teguritele, mis mõjutavad kondrotsüütide käitumist haigetes liigestes. Kuna aga liigesekõhre on aneuraalne ja avaskulaarne, ei ole see kude tõenäoliselt OA valu allikas. See asjaolu koos järeldustega, et liigesekõhre kahjustuse ja valu vahel puudub korrelatsioon OA patsientide [3,4, 5] või OA prekliiniliste mudelite [1] vahel, on põhjustanud fookuse nihke, et töötada välja ravimid tõhusaks valu kontrollimiseks. . Selles artiklis vaadeldakse liigesevalu uurimise uusimaid tulemusi ja tuuakse esile mõned esilekerkivad eesmärgid, mis võivad olla artriidi valuravi tulevikus (kokkuvõte joonisel XNUMX).
Tsütokiinide
Erinevate tsütokiinide toimed liigeste neurofüsioloogia uuringutes on viimasel ajal üsna silmapaistvalt esile tõstetud. Näiteks interleukiin-6 (IL-6) on tsütokiin, mis tavaliselt seondub membraaniga seotud IL-6 retseptoriga (IL-6R). IL-6 võib signaali anda ka lahustuva IL-6R-ga (SIL-6R) seondudes, et tekitada IL-6/sIL-6R kompleks. See IL-6/sIL-6R kompleks lübineerub seejärel transmembraanse glükoproteiini subühikuga 130 (gp130), võimaldades seeläbi IL-6-l anda signaali rakkudes, mis ei ekspresseeri konstitutiivselt membraaniga seotud IL-6R [25,26]. IL-6 ja SIL-6R on süsteemse põletiku ja artriidi võtmeosalised, kuna RA patsientide seerumis ja sünoviaalvedelikus on leitud mõlema ülesreguleerimist. [27,29]. Hiljuti täheldasid Vazquez jt, et IL-6/sIL-6R samaaegne manustamine roti põlvedele põhjustas põletikust põhjustatud valu, mis ilmnes seljaaju seljasarve neuronite reaktsiooni suurenemisest põlve ja teiste osade mehaanilisele stimulatsioonile. tagajäseme [30]. Seljaaju neuronite ülierutuvust täheldati ka siis, kui IL-6/sIL-6R kanti lokaalselt seljaajule. Lahustuva gp130 (mis eemaldaks IL-6/sIL-6R kompleksid, vähendades seeläbi trans-signalisatsiooni) spinaalne manustamine inhibeeris IL-6/sIL-6R-indutseeritud tsentraalset sensibilisatsiooni. Kuid lahustuva gp130 äge kasutamine üksi ei vähendanud neuronaalseid reaktsioone juba väljakujunenud liigesepõletikule.
Transient retseptorpotentsiaali (TRP) kanalid on mitteselektiivsed katioonkanalid, mis toimivad erinevate füsioloogiliste ja patofüsioloogiliste protsesside integraatoritena. Lisaks termosensatsioonile, kemosensatsioonile ja mehhanisatsioonile osalevad TRP kanalid valu ja põletiku moduleerimises. Näiteks on näidatud, et TRP vanilloid-1 (TRPV1) ioonikanalid aitavad kaasa liigesepõletikule, kuna termiline hüperalgeesia ei olnud TRPV1 monoartriidiga hiirtel esile kutsutud [31]. Sarnaselt on TRP anküriin-1 (TRPA1) ioonkanalid seotud artriidi mehhaanilise ülitundlikkusega, kuna retseptori blokeerimine selektiivsete antagonistidega nõrgendas mehaanilist valu Freundi täieliku adjuvantmudeli põletiku korral [32,33, 1]. Täiendavad tõendid selle kohta, et TRPV1 võib olla seotud OA valu neurotransmissiooniga, pärinevad uuringutest, milles OA naatriummonojodoatsetaadi mudelis on neuronaalse TRPV34 ekspressioon kõrgem [1]. Lisaks vähendas TRPV889425 antagonisti A-35 süsteemne manustamine monojodoatsetaadi mudelis lülisamba laia dünaamilise ulatuse ja notsitseptsioonispetsiifiliste neuronite esilekutsutud ja spontaanset aktiivsust [XNUMX]. Need andmed viitavad sellele, et endovanilloidid võivad olla seotud OA valuga seotud tsentraalsete sensibiliseerimisprotsessidega.
Praegu on teada, et TRPV1 kodeerivas geenis on vähemalt neli polümorfismi, mis põhjustavad ioonikanali struktuuri muutusi ja funktsiooni halvenemist. Üks konkreetne polümorfism (rs8065080) muudab TRPV1 tundlikkust kapsaitsiini suhtes ja seda polümorfismi kandvad isikud on termilise hüperalgeesia suhtes vähem tundlikud [36]. Hiljutises uuringus uuriti, kas rs8065080 polümorfismiga OA-patsiendid kogesid selle geneetilise anomaalia põhjal muutunud valutaju. Uurimisrühm leidis, et asümptomaatilise põlveliigese OA-ga patsientidel oli suurem tõenäosus rs8065080 geeni kandmiseks kui valulike liigestega patsientidel. [37]. See tähelepanek näitab, et OA patsiendid normaalse funktsioneerimisega; TRPV1 kanalitel on suurenenud liigesevalu risk ja see kinnitab veel kord TRPV1 võimalikku osalust OA valu tajumises.
Järeldus
Kuigi artriidi valu tõhusa ravi takistus püsib, on tehtud suuri hüppeid meie arusaamises neurofüsioloogilistest protsessidest, mis põhjustavad liigesevalu. Pidevalt avastatakse uusi sihtmärke, samal ajal kui teadaolevate radade taga olevaid mehhanisme määratletakse ja täpsustatakse. Ühe konkreetse retseptori või ioonikanali suunamine ei ole tõenäoliselt lahendus liigesevalu normaliseerimiseks, pigem on näidustatud polüfarmaatsia lähenemisviis, kus haiguse konkreetsetes faasides kasutatakse kombineeritult erinevaid vahendajaid. Funktsionaalsete ahelate lahtiharutamine valuraja igal tasandil parandab ka meie teadmisi liigesevalu tekke kohta. Näiteks liigesevalu perifeersete vahendajate tuvastamine võimaldab meil kontrollida liigeses toimuvat notsitseptsiooni ja tõenäoliselt vältida süsteemselt manustatavate farmakoteraapiate keskseid kõrvalmõjusid.
FAKTOGEENILINE PAIN
FACET SYNDROME & FACETOGENIC VALU
Facet'i sündroom on liigesehaigus, mis on seotud nimmepiirkonna liigeste ja nende innervatsiooniga ning tekitab nii lokaalset kui kiirgavat facetogeenset valu.
Lülisamba liigne pöörlemine, pikendamine või painutamine (korduv ülekoormus) võib põhjustada liigese kõhre degeneratiivseid muutusi. Lisaks võib see hõlmata degeneratiivseid muutusi teistes struktuurides, sealhulgas lülidevahelises kettas.
Emakakaela näo sündroom ja FACETOGENIC VALU
Aksiaalne kaelavalu (kiirgab harva üle õlgade), kõige sagedamini ühepoolselt.
Valu ja / või piirang laiendamine ja pöörlemine
Lõhna pärast palpeerumist
Hapnikuvastane valgus lokaalselt või õlgadele või ülemisele tagajärjele ning levib harva esiosa või alla käsivarre või sõrmedesse kui hernees ketas.
LUMBAR FACET SYNDROME & FACETOGENIC VALU
Valu või hellus alaseljas.
Kohalik hellus / jäikus alaselja kõrval.
Valu, jäikus või raskused teatud liigutustega (nt sirgelt püsti tõusmine või toolilt tõusmine).
Valu hüperekstensioonil
Eeldatav valu nimmepiirkonna ülaosa liigestest võib ulatuda kubemesse, puusasse ja reie ülemisse külgmisse ossa.
Eeldatav valu alumiste nimmeosa liigestest võib tungida sügavale reide, külgsuunas ja/või tagant.
L4-L5 ja L5-S1 tahkliigesed võivad viidata valule, mis ulatub distaalsesse külgmisse jalga ja harvadel juhtudel ka jalga.
Tõestatud meditsiin
Tõenduspõhine Interventsiooniparandus meditsiin vastavalt kliinilistele diagnoosidele
12. Valu, mis pärineb nimmepiirkonna liigestest
Abstraktne
Kuigi tahk-sündroomi olemasolu oli juba ammu kahtluse alla seatud, aktsepteeritakse seda nüüd kliinilise üksusena. Sõltuvalt diagnostilistest kriteeriumidest moodustavad zygapophysial liigesed kroonilise, aksiaalse alaseljavalu juhtudest vahemikus 5% kuni 15%. Kõige sagedamini tuleneb facetogeenne valu korduvast stressist ja/või kumulatiivsest madalast traumast, mis põhjustab liigesekapsli põletikku ja venitamist. Kõige sagedasem kaebus on aksiaalne alaseljavalu koos edasise valuga, mida tajutakse küljes, puusas ja reies. Ükski füüsilise läbivaatuse leid ei ole diagnoosimisel patognoomiline. Nimmepiirkonna facetogeense valu tugevaim indikaator on valu vähenemine pärast rami dorsales rami mediales (mediaalharud), mis innerveerivad tahke liigeseid. Kuna võib esineda valepositiivseid ja võib-olla ka valenegatiivseid tulemusi, tuleb tulemusi hoolikalt tõlgendada. Süstimisega kinnitatud sügapofüüsilise liigesevaluga patsientidel võib protseduurilisi sekkumisi läbi viia multidistsiplinaarse, multimodaalse raviskeemi raames, mis hõlmab farmakoteraapiat, füsioteraapiat ja regulaarset treeningut ning vajaduse korral psühhoteraapiat. Praegu on fasetogeense valu ravi kuldstandardiks raadiosagedusravi (1 B+). Intraartikulaarseid kortikosteroide toetavad tõendid on piiratud; seepärast tuleks see reserveerida neile, kes ei allu raadiosageduslikule ravile (2 B1).
Facetogeenne valu, mis tuleneb nimmepiirkonna liigestest, on täiskasvanud elanikkonna alaseljavalu tavaline põhjus. Goldthwaite kirjeldas sündroomi esimesena 1911. aastal ja üldiselt omistatakse Ghormleyle, kes lõi termini "fassettsündroom" aastal 1933. Facetogeenne valu on defineeritud kui valu, mis tuleneb mis tahes struktuurist, mis on osa tahkliigeste osast, sealhulgas kiudkapslist. , sünoviaalmembraan, hüaliinne kõhr ja luu.35
Sagedamini on see korduva stressi ja/või kumulatiivse madala trauma tagajärg. See põhjustab põletikku, mis võib põhjustada tahkliigese täitumist vedelikuga ja paisuda, mille tulemuseks on liigesekapsli venitamine ja sellele järgnev valu.27 Põletikulised muutused ümbermõõdetuna võivad ka seljaaju kitsendamise kaudu põhjustada seljaaju närvi, mis põhjustab ishiasi. Lisaks leidsid Igarashi jt., 28, et ventraalliinkapsli kaudu vabanevad põletikulised tsütokiinid sigapofüüsiaalses liigeste degeneratsiooniga patsientidel võivad osaliselt põhjustada seljaaju stenoosiga patsientide neuropaatilisi sümptomeid. Sigapofüüsiaalset liigesevalu põhjustavateks teguriteks on spondilolisteesia / lüüsi, degeneratiivse ketasti haigus ja vanus. 5
IC TÄIENDAVAD TESTID
Radioloogilisel uuringul esinevate tahkete liigeste patoloogiliste muutuste esinemissagedus sõltub katsealuste keskmisest vanusest, kasutatavast radioloogilisest tehnikast ja kõrvalekalde määratlusest. Degeneratiivseid tahke liigeseid saab kõige paremini visualiseerida arvutitulemuse (CT) uuringu abil. 49
NEUROPATILINE valu
Somatosensioonilises närvisüsteemis esmane kahjustus või düsfunktsioon on põhjustatud või põhjustatud valu.
Neuropaatilise valu on tavaliselt krooniline, raskesti ravitav ja sageli resistentne standardsele valuvaigistile.
Abstraktne
Neuropaatilist valu põhjustab somatosensoorse süsteemi kahjustus või haigus, sealhulgas perifeersed kiud (A-, A- ja C-kiud) ja tsentraalsed neuronid ning see mõjutab 7-10% kogu elanikkonnast. Kirjeldatud on mitmeid neuropaatilise valu põhjuseid. Tõenäoliselt suureneb selle esinemissagedus maailma elanikkonna vananemise, suhkurtõve sagenemise ja keemiaravi järgse vähi paranemise tõttu. Tõepoolest, tasakaalustamatus ergastava ja inhibeeriva somatosensoorse signaali vahel, muutused ioonikanalites ja varieeruvus selles, kuidas kesknärvisüsteemis valusõnumeid moduleeritakse, on kõik seotud neuropaatilise valuga. Lisaks näib kroonilise neuropaatilise valu koormus olevat seotud neuropaatiliste sümptomite keerukuse, halbade tulemuste ja raskete raviotsustega. Oluline on see, et neuropaatilise valuga patsientide elukvaliteet halveneb ravimite retseptide ja tervishoiuteenuste osutajate külastamise suurenemise ning haigestumuse tõttu valust endast ja esilekutsuvast haigusest. Vaatamata väljakutsetele soodustab neuropaatilise valu patofüsioloogia mõistmise areng uute diagnostiliste protseduuride ja isikupärastatud sekkumiste väljatöötamist, mis rõhutavad vajadust multidistsiplinaarse lähenemisviisi järele neuropaatilise valu ravis.
NEUROPATIINI PAINI PATOGENEESIS
PERIPHERILISED MEHHANISMIID
Pärast perifeerse närvi kahjustust muutuvad neuronid tundlikumaks ja neil tekib ebanormaalne erutuvus ja suurenenud tundlikkus stimulatsiooni suhtes.
Seda tuntakse ... perifeerne sensibiliseerimine!
KESKMECHANISMID
Perifeerias jätkuva spontaanse aktiivsuse tagajärjel areneb neuronite taustal suurenenud aktiivsus, laienevad vastuvõtlikud väljad ja suurenevad vastused aferentsetele impulssidele, sealhulgas normaalsetele taktiilsetele stiimulitele. Seda tuntakse ... Keskne sensibiliseerimine!
Krooniline neuropaatiline valu esineb sagedamini naistel (8% versus 5.7% meestel) ja üle 50-aastastel patsientidel (8.9% versus 5.6% alla 49-aastastel) ning see mõjutab kõige sagedamini alaselga ja alajäsemeid. , kael ja ülajäsemed24. Nimme- ja emakakaela valulikud radikulopaatid on ilmselt kõige sagedasem kroonilise neuropaatilise valu põhjus. Kooskõlas nende andmetega näitas Saksamaa valuspetsialistidele suunatud uuring >12,000 40 kroonilise valuga nii notsitseptiivse kui ka neuropaatilise valuga patsiendi seas, et 25% kõigist patsientidest koges vähemalt mõnda neuropaatilise valu tunnust (nt põletustunne, tuimus ja kipitus); kroonilise seljavalu ja radikulopaatiaga patsiendid olid eriti mõjutatudXNUMX.
Seni on pingetüüpi peavalu (TTH) kliinilisi neurofüsioloogilisi uuringuid läbi viidud kahe peamise eesmärgiga: (1) teha kindlaks, kas mõned neurofüsioloogilised parameetrid võivad toimida TTH markeritena, ja (2) uurida TTH füsiopatoloogiat. Seoses esimese punktiga on praegused tulemused pettumust valmistavad, kuna mõningaid TTH patsientidel leitud kõrvalekaldeid võib sageli täheldada ka migreenihaigetel. Teisest küljest on kliiniline neurofüsioloogia mänginud olulist rolli arutelus TTH patogeneesi üle. Ajutise lihase kontraktsiooni eksterotseptiivse pärssimise uuringud on tuvastanud ajutüve erutuvuse ja suprasegmentaalse kontrolli düsfunktsiooni. Sarnasele järeldusele jõuti ka trigeminocervikaalsete reflekside kasutamisel, mille kõrvalekalded TTH-s on viidanud ajutüve interneuronite inhibeeriva aktiivsuse vähenemisele, mis peegeldab ebanormaalseid endogeenseid valu kontrollimehhanisme. Huvitaval kombel näib TTH neuraalse erutuvuse anomaalia olevat üldine nähtus, mis ei piirdu ainult koljupiirkondadega. DNIC-sarnaste mehhanismide olemasolu on tõepoolest tõestatud ka somaatilistes piirkondades notsitseptiivse painderefleksi uuringutega. Kahjuks on enamikku TTH neurofüsioloogilisi uuringuid häirivad tõsised metodoloogilised vead, mida tuleks tulevastes uuringutes vältida, et TTH mehhanisme paremini selgitada.
viited:
Artriidi valu neurofüsioloogia. McDougall JJ1 Linton P.
Nimmepõhjas asuvate liigeste valu. van Kleef M1, Vanelderen P, Cohen SP, Lataster A, Van Zundert J, Mekhail N.
Neuropaatilise valuLuana Colloca,1Taylor Ludman,1Didier Bouhassira,2Ralf Baron,3Anthony H. Dickenson,4David Yarnitsky,5Roy Freeman,6Andrea Truini,7Nadine Attal, Nanna B. Finnerup,9Christopher Eccleston,1011Eija Kalso,12David L. Bennett,13Robert H. Dworkin,14ja Srinivasa N. Raja15
Kliinilise neurofüsioloogia panus pingetüüpi peavalu mehhanismide mõistmisse. Rossi P1, Vollono C, Valeriani M, Sandrini G.
Arstid määratlevad kroonilise valu, nagu mis tahes valu, mis kestab 3 kuni 6 kuud või rohkem. The valu mõjutab inimese vaimset tervist ja igapäevaelu. Valu pärineb mitmest sõnumist, mis läbib närvisüsteemi. Näib, et depressioon tekitab valu. See põhjustab tõsiseid sümptomeid, mis mõjutavad seda, kuidas inimene tunneb, mõtleb ja kuidas igapäevaseid tegevusi, st magamine, söömine ja töötamine. Chiropractor Dr. Alex Jimenez uhkeldab potentsiaalsetest biomarkeritest, mis võivad aidata leida ja ravida valu ja kroonilise valu algpõhjuseid.
Edukate valude juhtimise esimene samm on terviklik biopsiotsühhosotsiaalne hinnang.
Orgaanilise patoloogia ulatus ei pruugi valulikkuses õigesti kajastada.
Esialgset hinnangut saab kasutada selliste valdkondade kindlakstegemiseks, mis nõuavad põhjalikumat hindamist.
Kroonilise valu mõju hindamiseks on kättesaadavad paljud valideeritud enesearvestusvahendid.
Kroonilise valu hindamine
Krooniline valu on rahvatervisega seotud probleem, mis mõjutab 20-30% lääneriikide elanikkonnast. Kuigi valu neurofüsioloogia mõistmisel on olnud palju teaduslikke edusamme, ei ole patsiendi kroonilise valu probleemi täpselt hinnata ja diagnoosida otseselt ega täpselt määratletud. Kroonilise valu kujundamine mõjutab seda, kuidas valu hinnatakse ja kroonilise valu diagnoosimisel arvestatakse tegureid. Orgaanilise patoloogia ja valu intensiivsuse suuruse või tüübi vahel ei ole üks-ühele seost, vaid selle asemel, kroonilise valu kogemus on kujundanud hulgaliselt biomeditsiiniliste, psühhosotsiaalsete (nt patsientide uskumused, ootused ja meeleolu) ja käitumuslikke tegureid (nt seoses vastuseid teistele olulistele inimestele). Kroonilise valuga isiku igakülgse hindamise hindamine kummaski neist valdkondadest on oluline raviotsuste tegemiseks ja optimaalsete tulemuste hõlbustamiseks. See hindamine peaks sisaldama põhjalikku patsiendi ajalugu ja meditsiinilist hinnangut ning lühikest sõeluuringut, kus saab jälgida patsiendi käitumist. Esialgse hindamise käigus tuvastatud küsimuste edasine hindamine juhib otsuseid selle kohta, millised täiendavad hinnangud, kui neid on, võivad olla asjakohased. Standardiseeritud ise aru vahendeid hindama patsiendi valu intensiivsust, funktsionaalne võimeid, uskumusi ja ootusi ning emotsionaalne pinge on olemas, ja võib manustada arsti või esildise põhjalikumat hindamist saab aidata ravi planeerimisel.
Valu on väga levinud sümptom. Hinnanguliselt mõjutab krooniline valu ainult 30% USA täiskasvanud elanikkonnast, ulatudes 100 miljonit täiskasvanutest. 1
Vaatamata kroonilise valuga inimesi ravitava hinna suurenemisele, on paljudel patsientidel esinenud leevendust raske ja valu täielik kõrvaldamine on haruldane. Ehkki teadmised valu neurofüsioloogia kohta on olnud märkimisväärsed, koos tugevate valuvaigistavate ravimite väljatöötamise ning muude uuenduslike meditsiiniliste ja kirurgiliste sekkumistega on valude vähendamine olemasolevate protseduuride abil keskmiselt 30 40% ja see juhtub vähem kui pooled ravitud patsientidest.
Valu mõtlemine mõjutab seda, kuidas me valu hindame. Hindamine algab ajaloo ja füüsilise uurimisega, millele järgneb laboratoorsete analüüside ja diagnostilise pildistamise protseduur, et tuvastada ja / või kinnitada mistahes sümptomit / s või selle põhjustanud patoloogiat valu generaator.
Identifitseeritava orgaanilise patoloogia puudumisel võib tervishoiuteenuse osutaja eeldada, et sümptomite aruanne tuleneb psühholoogilistest teguritest ja võib nõuda psühholoogilist hindamist patsiendi aruande aluseks olevate emotsionaalsete tegurite avastamiseks. Sümptomite aruanne on kahesuguse iseloomuga somaatiline or psühhogeensed mehhanismid.
Näiteks orgaanilised alused mõnede kõige levinumate ja korduvate akuutsete (nt peavalu) 3 ja krooniliste [nt seljavalu, fibromüalgia (FM)] valu probleemid on suures osas teadmata, 4,5 aga teistel juhtudel võivad asümptomaatilised indiviididel esineda struktuurseid kõrvalekaldeid nagu hernide kettad, mis selgitavad valu, kui see esineb. 6,7Puuduvad piisavad selgitused tuvastamata orgaanilise patoloogiaga patsientidele, kes teatavad tugevast valust ja valuvabadest inimestest, kellel on märkimisväärne ja objektiivne patoloogia.
Krooniline valu mõjutab enamasti ainult üksikut patsienti, vaid ka tema olulisi teisi (partnerid, sugulased, tööandjad ja kaaskodanikud ja sõbrad), mis muudab asjakohase ravi oluliseks. Rahuldav ravi võib tulla üksnes põhjaliku hinnangu bioloogilise etioloogia valu koos patsiendi spetsiifilise psühhosotsiaalse ja käitumuslike esitlus, sealhulgas nende emotsionaalset seisundit (nt ärevus, depressioon ja viha), taju ja mõistmist sümptomid ja reaktsioonid nendele sümptomid märkimisväärsete teiste poolt. 8,9 Peamine eeldus on see, et mitmed tegurid mõjutavad kroonilise valu sümptomeid ja funktsionaalseid piiranguid. Seepärast on vaja terviklikku hinnangut, mis käsitleb biomeditsiini, psühhosotsiaalseid ja käitumuslikke valdkondi, kuna kõik aitavad kaasa kroonilisele valule ja sellega seotud puudele. 10,11
Kroonilise valuga isiku terviklik hindamine
Türk ja Meichenbaum12 soovitas suunata inimesi, kes valule teatavad, hinnata kolme keskset küsimust:
Milline on patsiendi haiguse või vigastuse ulatus (füüsiline nõrkus)?
Mis on haiguse suurus? See tähendab, mil määral patsient kannatab, on puudega ja ei suuda tavalist tegevust nautida?
Kas üksikisiku käitumist ilmselt asjakohane haiguse või vigastuse või on olemas tõendid sümptom võimendus tahes erinevaid psühholoogilised või sotsiaalsed põhjused (nt kasu nagu positiivse tähelepanu, tujumuutvad ravimeid, rahalist hüvitist)?
Nendele küsimustele vastamiseks tuleks patsiendilt koguda teavet anamneesi ja füüsilise läbivaatuse järgi koos kliinilise intervjuuga ja standardiseeritud hindamisvahendite abil. Tervishoiuteenuse osutajad peavad füüsilise läbivaatuse ja diagnostiliste testide abil otsima mis tahes valu põhjuse (d), hinnates samaaegselt patsiendi meeleolu, hirme, ootusi, toimetulekupingutusi, ressursse, oluliste teiste reaktsioone ja valu mõju patsientidele. elab.11 Lühidalt öeldes peab tervishoiuteenuse osutaja hindama mitte ainult valu, vaid tervet inimest.
Ajaloo ja meditsiinilise hindamise üldised eesmärgid on:
(i) täiendava diagnostilise testimise vajalikkust
(ii) määrama kindlaks, kas meditsiinilised andmed võivad selgitada patsiendi sümptomeid, sümptomite raskust ja funktsionaalseid piiranguid
(iii) teha meditsiiniline diagnoos
(iv) hinnata sobiva ravi kättesaadavust
v) kehtestada ravi eesmärgid
(vi) määrab sobiva kursuse sümptomite juhtimiseks, kui täielik ravi ei ole võimalik.
Märkimisväärne hulk patsiente, kes teatavad kroonilisest valust, ei näita füüsilist patoloogiat, kasutades tavalisi radiograafi, arvutatud teljmotograafia skaneeringuid või elektromüograafiat (laialdane kirjandus on kättesaadav füüsilise hindamise, radiograafia ja laboratoorse hindamise protseduuride kohta valu füüsilise aluse kindlakstegemiseks), 17 teeb täpse patoloogilise diagnoosi keeruliseks või võimatuks.
Hoolimata nendest piirangutest, patsiendi anamneesi ja läbivaatuse aluseks jääma meditsiinilise diagnoosi, võib pakkuda kaitsemehhanismi üle tõlgendamisel leide diagnostiliste, mis on suures osas kinnitava, ning seda saab kasutada, et suunata suunas edasist hindamist jõupingutusi.
Lisaks pakuvad kroonilise valu probleemidega patsiendid sageli erinevaid ravimeid.18 On oluline arutada patsiendi praeguseid ravimeid intervjuu ajal, kuna paljud valuvaigistid on seotud kõrvalmõjudega, mis võivad põhjustada või jäljendada emotsionaalset stressi. 19 Tervishoiuteenuste osutajad peaksid teadma mitte ainult kroonilise valu kasutatavaid ravimeid, vaid ka selliste ravimite kõrvaltoimeid, mis põhjustavad väsimust, unehäireid ja meeleolu muutusi, et vältida depressiooni väärdiagnoosimist.
Igapäevaste päevikute kasutamine on arvatavasti täpsem, kuna need põhinevad pigem reaalajas kui tagasivõtmisel. Patsientidelt võidakse paluda regulaarselt säilitada valu intensiivsusega päevikuid, mis registreeritakse mitu korda päevas (nt söögikordus ja magamaminek) mitu päeva või nädalat registreeritavate reitingutega ning mitu korda saab valu hinnata.
Üks paber- ja pliiatsipäevikute kasutamisel täheldatud probleem on see, et patsiendid ei pruugi järgida juhiseid reitingute andmiseks kindlaksmääratud intervallidega. Pigem võivad patsiendid päevikuid koostada eelnevalt („täitke edasi“) või vahetult enne arsti poole pöördumist („täitke tagurpidi“), 24 õõnestades päevikute oletatavat kehtivust. Elektroonilised päevikud on nende probleemide vältimiseks mõnes uurimuses leidnud heakskiitu.
Uuringud on näidanud tervisega seotud elukvaliteedi (HRQOL) hindamise olulisust kroonilise valuga patsientidel lisaks funktsionaalrühmale. 31,32 On olemas mitmeid väljakujunenud, psühhomeetriliselt toetatud HRQOL-i meetmeid [Medical Outcome Studies Short-Form Health Survey (SF 36)], 33-i üldised füüsilise funktsioneerimise näitajad (nt Pain Disability Index (PDI)), 34 ja haigusepõhised meetmed [nt Lääne-Ontario MacMasteri osteoartriidi indeks (WOMAC); 35 Roland-Morrise seljavalu keeleküsitluse küsimustik (RDQ)]; 36 funktsiooni ja elukvaliteedi hindamiseks.
Haiguspetsiifilised meetmed on kavandatud selleks, et hinnata konkreetse seisundi (nt valu ja jäikus osteoartriidiga inimestel) mõju, samas kui üldised meetmed võimaldavad võrrelda antud haigusega seotud füüsilist toimet ja selle mitmesuguste muude haigusseisundite ravimist. Üldise meetme kasutamisel ei pruugi haiguse spetsiifilisi mõjusid tuvastada; seetõttu võivad haigusepõhised meetmed kliiniliselt oluliselt parandada või halvendada spetsiifilisi funktsioone ravi tulemusena. Toimimise üldised mõõtmised võivad osutuda kasulikuks, kui võrrelda paljude valulike seisunditega patsiente. Haiguspetsiifiliste ja üldiste meetmete kombineeritud kasutamine hõlbustab mõlema eesmärgi saavutamist.
Emotsionaalse stressi olemasolu kroonilise valu inimestel tekitab väljakutse selliste sümptomite hindamisel nagu väsimus, aktiivsuse vähenemine, libiido vähenemine, söögiisu muutused, unehäired, kehakaalu suurenemine või kadu ning mälu ja kontsentratsiooni defitsiit, kuna need sümptomid võivad olla valu, emotsionaalse distressi või valu raviks ette nähtud ravimite tulemus.
Spetsiaalselt valupatsientidele on välja töötatud instrumendid, et hinnata psühholoogilist stressi, valu mõju patsientide elule, kontrollitunnet, toimetulekukäitumist ning suhtumist haigustesse, valu ja tervishoiuteenuse osutajatesse.
Näiteks Beck Depressiooni Inventari (BDI) 39 ja Mood State Profiil (POMS) 40 on psühhomeetriliselt usaldusväärsed, et hinnata depressiooniga meeleolu, emotsionaalse stressi ja meeleoluhäirete sümptomeid ning on soovitatav kasutada kõigis kliinilistes uuringutes krooniline valu; 41 aga tuleb skoori tõlgendada ettevaatusega ning võib osutuda vajalikuks muuta emotsionaalse distressi taseme kriteeriume, et vältida valepositiivseid tulemusi. 42
Lab biomarkerid valu
Biomarkerid on bioloogilised omadused, mida saab kasutada tervise või haiguse märkimiseks. Käesolevas paberkandjal käsitletakse uuringuid madalama seljavalu (LBP) biomarkeritega inimestel. LBP on puuete peamine põhjus, mis on tingitud mitmest lülisambaga seotud häiretest, sh perterosiilsete ketaste degeneratsioon, ketta hernikumine, seljaaju stenoos ja nägatõbi. Nendes uuringutes keskendutakse põletikulistele mediaatoritele, sest põletik aitab kaasa plaadi degeneratsiooni ja sellega seotud valu mehhanismide patogeneesile. Uuringud näitavad üha sagedamini, et põletikuliste mediaatorite esinemist saab süsteemis veres mõõta. Need biomarkerid võivad olla uudsed vahendid patsiendi ravi suunamiseks. Praegu on patsiendi reaktsioon ravile ettearvamatu, kui see on korduvalt esinenud, ja kui kirurgiline ravi võib anda anatoomilise korrigeerimise ja valu leevenduse, on nad invasiivsed ja kulukad. Läbivaatamine hõlmab uuringuid, mis on tehtud populatsioonidega, kellel on spetsiifiline diagnoos ja LBP määratlemata päritolu. Kuna LBP loodusajalugu on progresseeruv, liigitatakse uuringute ajaline olemus sümptomoloogia / haiguse kestusega. Vaadatakse läbi ka uuringud ravimisega seotud biomarkeritega toimunud muutuste kohta. Lõppkokkuvõttes on LBP diagnoosimis biomarkerid ja seljaaju degenereerumine potentsiaalselt varjatud individuaalse lülisambarantiini ajastu isikupärastatud raviks LBP ravis.
Kroonilise neuropaatilise valu biomarkerid ja potentsiaalne rakendamine seljaaju stimuleerimisel
Selles ülevaates keskenduti arusaamale, millised ained inimese kehas kasvavad ja vähenevad neuropaatilise valu suurenemisega. Vaatasime läbi mitmesugused uuringud ja nägime korrelatsioone neuropaatilise valu ja immuunsussüsteemi komponentide vahel (see süsteem kaitseb keha haiguste ja infektsioonide eest). Meie leiud on eriti kasulikud selleks, et mõista ebamugavuse vähendamise või kõrvaldamise viise, mille tagajärjel tekib krooniline neuropaatiline valu. Seljaaju stimulatsiooni (SCS) protseduur on üks väheseid tõhusaid parandavaid valu ravimeetodeid. Järelevalve uuringus rakendatakse meie läbivaatamisel tehtud järeldusi SCSile, et mõista mehhanismi ja tõhusust veelgi optimeerida.
On leitud, et põletikuvastased tsütokiinid nagu IL-1a, IL-6, IL-2, IL-33, CCL3, CXCL1, CCR5 ja TNF-a mängivad olulist rolli krooniliste valude seisundite amplifitseerimisel.
Pärast erinevate valu biomarkeritega seotud uuringute ülevaatamist leidsime, et põletikueelsete tsütokiinide ja kemokiinide, nagu IL-1a, IL-6, IL-2, IL-33, CCL3, CXCL1, CCR5 ja TNF, tase seerumis -? olid kroonilise valu ajal oluliselt reguleeritud. Teiselt poolt leiti, et põletikuvastased tsütokiinid, nagu IL-10 ja IL-4, näitavad kroonilise valuseisundi ajal märkimisväärset allaregulatsiooni.
Depressiooni biomarkerid
Paljude uuringutega on kaasatud sadu eeldatavaid depressiooni biomarkereid, kuid ei ole veel täielikult välja selgitanud oma rolli depressiivses haiguses või leidnud, milline on ebanormaalne, millistel patsientidel ja kuidas bioloogilist teavet saab kasutada diagnoosi, ravi ja prognoosi parandamiseks. Selline edusammude puudumine on osaliselt tingitud depressiooni olemusest ja heterogeensusest koos teadusliku kirjanduse metodoloogilise heterogeensusega ja potentsiaaliga biomarkerite suur hulgaga, mille ekspressioon muutub sageli erinevate tegurite järgi. Vaatame olemasolevat kirjandust, mis näitab, et põletikuliste, neurotroofsete ja ainevahetusprotsesside, samuti neurotransmitterite ja neuroendokriinsüsteemi komponentidega seotud markerid on väga paljutõotavad kandidaadid. Neid võib mõõta geneetiliste ja epigeneetiliste, transkriptoomsete ja proteoomiliste, metabolomiliste ja neuroimaging hindamiste abil. Uute lähenemisviiside ja süstemaatiliste uurimisprogrammide kasutamine on nüüd vajalik, et teha kindlaks, kas ja milliseid biomarkereid saab ravile reageerimise prognoosimiseks kasutada, stratifitseerige patsiendid spetsiifiliste ravimitega ja välja töötada eesmärgid uute sekkumiste jaoks. Me järeldame, et nende teadustööde edasiarendamise ja laiendamise kaudu on palju lubadusi depressiooni koormuse vähendamiseks.
viited:
Kroonilise valuga patsientide hindamineEJ Dansiet ja DC Turk * t
Valu on inimese keha loomulik vastus vigastusele või haigusele ja see on sageli hoiatus, et midagi on valesti. Kui probleem on paranenud, peame üldiselt selle valulikke sümptomeid kogema, aga mis juhtub, kui valu jätkub kaua pärast põhjuse kadumist? Krooniline valu on meditsiiniliselt defineeritud kui püsiv valu, mis kestab 3 kuni 6 kuud või rohkem. Krooniline valu on kindlasti keeruline elamine, mis mõjutab kõike üksikisiku aktiivsuse tasemest ja töövõimest ning isiklikest suhetest ja psühholoogilistest seisunditest. Kuid kas teate, et krooniline valu võib samuti mõjutada teie aju struktuuri ja funktsioone? Selgub, et need aju muutused võivad põhjustada nii kognitiivseid kui ka psühholoogilisi häireid.
Krooniline valu ei mõjuta ainult mõnda eripiirkonda, sest tegelikult võib see põhjustada muutusi paljudes olulistes aju valdkondades, millest enamus on seotud paljude põhiprotsesside ja -funktsioonidega. Aastate jooksul on mitmesuguseid uurimusi läbi vaadatud muutused hipokampuses koos halli aine redutseerumisega dorsolateral prefrontaalsest korteksist, amygdalast, ajutüvest ja parema isoleeritud koorega, nimetades vaid mõnda, mis on seotud kroonilise valuga. Mõne nendest piirkondadest koosneva struktuuri ja nendega seotud funktsioonide jagunemine võib aidata kaasa nende aju muutustele kontekstis paljude kroonilise valu puhul. Järgmise artikli eesmärk on näidata ja arutada kroonilise valuga seotud struktuurseid ja funktsionaalseid aju muutusi, eriti juhul, kui need kajastavad tõenäoliselt mitte kahju ega atroofiat.
Aju muutused kroonilises valu ei mõjuta tõenäoliselt kahjustusi ega atroofiat
Abstraktne
Krooniline valu näib olevat seotud aju halli aine vähenemisega piirkondades, mis on tingitud valu edasikandumisest. Nende struktuurimuutuste aluseks olevad morfoloogilised protsessid, tõenäoliselt pärast funktsionaalseid ümberkorraldusi ja aju tsentraalset plastilisust, jäävad ebaselgeks. Puusaliigese artroosi valu on üks väheseid kroonilisi valusündroome, mis on peamiselt ravitavad. Uurisime 20 patsienti, kellel oli krooniline valu ühepoolse koksartroosi tõttu (keskmine vanus 63.25-9.46 (SD) aastat, 10 naist) enne puusaliigese endoproteesimist (valu seisund) ja jälgisime aju struktuurimuutusi kuni 1 aasta pärast operatsiooni: 6 nädalat , 8 nädalat ja 12 kuud, kui valu on täielikult vaba. Patsientidel, kellel oli ühepoolse koksartroosi tagajärjel tekkinud krooniline valu, oli halli ainet oluliselt vähem, võrreldes kontrollidega eesmise tsingulaarkoore (ACC), isolaarkoore ja operkuli, dorsolateraalse prefrontaalse koore (DLPFC) ja orbitofrontaalse korteksi korral. Need piirkonnad toimivad kogemuste ja valu ennetamise ajal multiintegratiivsete struktuuridena. Kui patsiendid pärast endoproteetilistest operatsioonidest taastumist valusid, leiti halli aine suurenemine peaaegu samades piirkondades. Samuti leidsime aju halli aine progresseeruva kasvu premotoorses ajukoores ja täiendavas motoorses piirkonnas (SMA). Me jõuame järeldusele, et kroonilise valu hallide ainete kõrvalekalded ei ole haiguse põhjus, vaid sekundaarsed ning on vähemalt osaliselt tingitud motoorsete funktsioonide ja keha integreerumise muutustest.
Sissejuhatus
Kroonilise valuga patsientide funktsionaalse ja struktuurse ümberkorraldamise tõendid toetavad ideed, et kroonilist valu ei tuleks kontseptualiseerida üksnes muutunud funktsionaalse seisundina, vaid ka aju funktsionaalse ja struktuurse plastilisuse tagajärjena [1], [2], [3], [4], [5], [6]. Viimase kuue aasta jooksul on avaldatud üle 20 uuringu, mis tõestavad aju struktuurseid muutusi 14 kroonilise valu sündroomi korral. Kõigi nende uuringute silmatorkav omadus on asjaolu, et halli aine muutused ei jaotunud juhuslikult, vaid toimusid määratletud ja funktsionaalselt väga spetsiifilistes ajupiirkondades, nimelt osalemine supraspinaalses notsitseptiivses protsessis. Silmapaistvamad leiud olid iga valusündroomi puhul erinevad, kuid kattusid tsingulaarkoores, orbitofrontaalses ajukoores, insula ja seljaosas. [4] Edasised struktuurid hõlmavad taalamust, dorsolateraalset prefrontaalset ajukooret, basaalganglione ja hipokampuse piirkonda. Neid järeldusi käsitletakse sageli raku atroofiana, mis tugevdab aju halli aine kahjustuse või kaotuse ideed [7], [8], [9]. Tegelikult leidsid teadlased korrelatsiooni aju halli aine vähenemise ja valu kestuse vahel [6], [10]. Kuid valu kestus on seotud ka patsiendi vanusega ning vanusest sõltuv halli aine globaalne, aga ka piirkondlikult spetsiifiline langus on hästi dokumenteeritud [11]. Teiselt poolt võivad need struktuurimuutused olla ka raku suuruse vähenemine, rakuvälised vedelikud, sünaptogenees, angiogenees või isegi veremahu muutustest [4], [12], [13]. Ükskõik, mis allikas see on, on selliste leidude tõlgendamiseks oluline näha neid morfomeetrilisi leide paljude morfomeetriliste uuringute valguses treeningust sõltuvast plastilisusest, arvestades, et piirkondlikult spetsiifilisi aju struktuurseid muutusi on kognitiivsete ja füüsiliste harjutuste järel korduvalt näidatud [ 14].
Pole arusaadav, miks ainult suhteliselt väikesel osal inimestest tekib krooniline valusündroom, arvestades, et valu on universaalne kogemus. Tekib küsimus, kas mõnel inimesel võib keskvalu edastavate süsteemide struktuurne erinevus toimida kroonilise valu diateesina. Amputatsioonist [15] ja seljaaju vigastusest [3] tulenevad halli aine muutused fantoomvalus näitavad, et aju morfoloogilised muutused on vähemalt osaliselt kroonilise valu tagajärg. Kuid puusaliigese artroosi (OA) valu on üks väheseid kroonilise valu sündroomi, mis on peamiselt ravitav, kuna 88% neist patsientidest on puusaliigese täieliku asendamise (THR) operatsiooni järgselt regulaarselt valudeta [16]. Pilootuuringus oleme analüüsinud kümmet puusa OA-ga patsienti enne ja vahetult pärast operatsiooni. Enne THR-i operatsiooni leidsime kroonilise valu ajal halli aine vähenemist eesmises tsingulaarses ajukoores (ACC) ja isolaadis ning leidsime pärast operatsiooni valuvaba seisundi halli aine suurenemist vastavatel ajupiirkondadel [17]. Sellele tulemusele keskendudes laiendasime nüüd oma uuringuid, milles uuriti rohkem patsiente (n? =? 20) pärast edukat THR-i ja jälgisime aju struktuurseid muutusi nelja ajaintervalliga kuni ühe aasta jooksul pärast operatsiooni. Motoorikast paranemisest või depressioonist tingitud halli aine muutuste kontrollimiseks viisime läbi ka motoorse funktsiooni ja vaimse tervise parandamise küsimustikud.
Materjalid ja meetodid
Vabatahtlikud
Siin teatatud patsiendid on hiljuti avaldatud 20 patsiendist 32 patsiendi alarühm, keda võrreldi vanuse ja soo järgi sobiva tervisliku kontrollrühmaga [17], kuid osalesid täiendavas üheaastases järelkontrollis. Pärast operatsiooni langes 12 patsienti teise endoproteetilise operatsiooni (n = = 2), raske haiguse (n = = 2) ja nõusoleku tühistamise (n = = 8) tõttu. See jättis kahekümnest patsiendist koosneva ühepoolse puusaliigese OA-ga (keskmine vanus 63.25-9.46 (SD) aastat, 10 naist), keda uuriti neli korda: enne operatsiooni (valu seisund) ja uuesti 6 ja 8 nädalat ning 12 18 kuud pärast endoproteetilist operatsiooni, kui see on täielikult valuvaba. Kõigil esmase puusaliigese OA-ga patsientidel oli valu anamnees üle 10 kuu, ulatudes 14 kuni 12 aastani (keskmine 1 aastat) ja keskmine valu skoor 33 (vahemikus 7.35 kuni 65.5) visuaalse analoogkaala (VAS) vahemikus 40 (valu pole) kuni 90 (halvim, mida on võimalik ette kujutada). Hinnasime väiksemate valusündmuste, sealhulgas hamba-, kõrva- ja peavalude esinemist kuni 0 nädalat enne uuringut. Valisime juhuslikult ka andmed ülalnimetatud pilootuuringu 100-st 4-st soost ja vanusest vastanud tervislikest kontrollgruppidest (keskmine vanus 20-60,95 (SD) aastat, 8,52 naist) [10]. Ühelgi 32-st patsiendist ega 17-st sugu- ja vanuserühmas tervetest vabatahtlikest ei olnud neuroloogilisi ega sisehaigusi. Kohalik eetikakomitee andis uuringule eetilise heakskiidu ja kõigilt uuringus osalejatelt saadi enne uuringut kirjalik nõusolek.
Käitumuslikud andmed
Kogusime andmeid kõigi patsientide depressiooni, somatiseerimise, ärevuse, valu ning füüsilise ja vaimse tervise kohta ning kõigi nelja ajapunkti kohta, kasutades järgmisi standardiseeritud küsimustikke: Becki depressiooni inventuur (BDI) [18], Brief Symptom Inventory (BSI) [19], Schmerzempfindungs-Skala (SES? =? Valu ebameeldivuste skaala) [20] ja terviseküsitluse 36 elemendi lühivorm (SF-36) [21] ja Nottinghami terviseprofiil (NHP). Viisime läbi korduvaid meetodeid ANOVA ja sidusime kahesabilised t-testid, et analüüsida käitumise pikisuunalisi andmeid, kasutades Windowsi SPSS 13.0 (SPSS Inc., Chicago, IL), ja sfäärilisuse eelduse rikkumise korral kasutasime Greenhouse Geisseri parandust. Olulisuse tasemeks määrati p <0.05.
VBM - andmete hankimine
Kujutise omandamine. Kõrge eraldusvõimega MR-skaneerimine viidi läbi 3T MRI-süsteemil (Siemens Trio) tavalise 12-kanalilise peamähisega. Kõigi nelja ajapunkti puhul skaneerige I (1–3 kuud enne endoproteetilist operatsiooni), II skaneerimine (6–8 nädalat pärast operatsiooni), III skaneerimine (12–18 nädalat pärast operatsiooni) ja IV skaneerimine (10) kuud pärast operatsiooni), igale patsiendile tehti T14 kaalutud struktuurne MRI, kasutades 1D-FLASH järjestust (TR 3 ms, TE 15 ms, pöördenurk 4.9, 25 mm viilud, FOV 1-256, voksli suurus 256 1 mm).
Pilditöötlus ja statistiline analüüs
Andmete eeltöötlus ja analüüs viidi läbi SPM2-ga (Wellcome Department of Cognitive Neurology, London, Suurbritannia), mis töötas Matlabi all (Mathworks, Sherborn, MA, USA) ja sisaldas vokselipõhist morfomeetriat (VBM) - pikisuunaliste andmete tööriistakasti. põhineb kõrge eraldusvõimega struktuurilistel 3D MR-piltidel ja võimaldab rakendada voxel-statistikat, et tuvastada halli aine tiheduse või mahtude piirkondlikke erinevusi [22], [23]. Eeltöötlus hõlmas kokkuvõtlikult ruumilist normaliseerimist, halli aine segmenteerimist ja 10 mm ruumilist silumist Gaussi tuumaga. Eeltöötluse etappide jaoks kasutasime optimeeritud protokolli [22], [23] ning skanneri ja uuringu spetsiifilist halli aine malli [17]. Selle analüüsi meie pilootuuringuga võrreldavaks muutmiseks kasutasime SPM2, mitte SPM5 või SPM8 [17]. kuna see võimaldab pikisuunalisi andmeid suurepäraselt normaliseerida ja segmenteerida. Kuid kuna hiljuti sai kättesaadavaks VBM-i uuem värskendus (VBM8) (dbm.neuro.uni-jena.de/vbm/), kasutasime ka VBM8.
Cross-Sectional analüüs
Aju halli aine piirkondlike erinevuste tuvastamiseks rühmade vahel (patsiendid aja I skaneerimisel (krooniline valu) ja tervislik kontroll) kasutasime kaheproovilist t-testi. Rakendasime kogu aju künniseks p <0.001 (parandamata) meie tugeva põhihüpoteesi tõttu, mis põhineb 9 sõltumatul uuringul ja kohordil, mis näitavad halli aine vähenemist kroonilise valuga patsientidel [7], [8], [ 9], [15], [24], [25], [26], [27], [28], et halli aine suurenemine ilmneb samades (valutöötlemiseks asjakohastes) piirkondades nagu meie pilootuuringus (17). ). Rühmad sobitati vanuse ja soo järgi, ilma et rühmade vahel oleks olulisi erinevusi. Et uurida, kas rühmade vahelised erinevused ühe aasta pärast muutusid, võrdlesime patsiente ka IV aja skaneerimisel (valuvaba, üks aasta jälgimine) meie tervisliku kontrollrühmaga.
Pikisuunaline analüüs
Ajapunktide erinevuste tuvastamiseks (Scan IIV) võrreldi skaneeringuid enne operatsiooni (valu seisund) ja uuesti 6 ja 8 nädalat ning 12 kuud pärast endoproteetilist operatsiooni (valuvaba) ANOVA kordusmeetmena. Kuna kõik kroonilise valu tõttu tekkinud aju muutused võivad pärast operatsiooni ja valu peatumist vajada veidi aega ja operatsioonijärgse valu tõttu, millest patsiendid teatasid, võrdlesime pikisuunalises analüüsis I ja II skaneeringut III ja IV skaneerimisega. Muudatuste tuvastamiseks, mis pole valuga tihedalt seotud, otsisime ka progressiivseid muutusi kõigi ajaintervallide jooksul. Pöörasime vasaku puusa OA-ga patsientide aju (n? =? 18), et normaliseerida nii valu kui ka gruppide võrdluse ja pikisuunalise analüüsi jaoks, kuid analüüsisime ennekõike ümberlükatud andmeid. BDI skoori kasutasime mudelis kovariaadina.
Tulemused
Käitumuslikud andmed
Kõik patsiendid teatasid enne operatsiooni kroonilisest puusavalust ja olid vahetult pärast operatsiooni valud (seoses selle kroonilise valuga), kuid teatasid II skaneerimisel üsna ägedast operatsioonijärgsest valust, mis erines osteoartriidist tingitud valust. Vaimse tervise skoor SF-36 (F (1.925 / 17.322) p = 0.352, p? = 0.7) ja BSI globaalse skoori GSI (F (1.706 / 27.302)? = 3.189. 0.064, p? = XNUMX XNUMX ) ei näidanud aja jooksul muutusi ega vaimset kaasuvat haigestumist. Ükski kontrollgrupp ei teatanud ägedast ega kroonilisest valust ning ühelgi ei olnud depressiooni ega füüsilise / vaimse puude sümptomeid.
Enne operatsiooni ilmnes mõnel patsiendil BDI skooris kerge kuni mõõdukas depressioonisümptomaatika, mis oluliselt langes III skaneerimisel (t (17) a = 2.317 0.033, p? = 16 2.132) ja IV (t (0.049) p = 16. 4.676, p? =? 0.001). Lisaks paranesid kõigi patsientide SES-skoorid (valu ebameeldivused) märkimisväärselt alates I skaneerimisest (enne operatsiooni) kuni II skannimiseni (t (14) a =? 4.760, p <0.001), III skaneeringuga (t (14) a =? 4.981, p <0.001) ja skaneerimine IV (t (1) a = 1. 2, p <3, 4 aasta pärast operatsiooni), kuna valu ebameeldivus vähenes valu intensiivsusega. Valu hinnang 1. ja 2. skaneerimisel olid positiivsed, sama hinne 19.6. ja 1. päeval. SES kirjeldab ainult tajutava valu kvaliteeti. Seetõttu oli see 13.5. ja 2. päeval positiivne (3. päeval keskmiselt 4 ja XNUMX. päeval XNUMX) ning XNUMX. ja XNUMX. päeval negatiivne (na). Kuid mõned patsiendid ei mõistnud seda protseduuri ja kasutasid SES-i globaalse kvaliteediga elu mõõt. Seetõttu küsiti kõigilt patsientidelt valu esinemise kohta samal päeval individuaalselt ja sama isiku poolt.
Lühivormis terviseküsitlus (SF-36), mis koosneb füüsilise tervise skoori ja vaimse tervise skoori kokkuvõtlikest meetmetest [29], paranesid patsiendid füüsilise tervise skooris I skaneerimisest II skannini (t ( 17) p = 4.266, p = = 0.001), skaneerimine III (t (16) p = 8.584, p <0.001) ja IV (t (12) p = 7.148, p <0.001), kuid mitte vaimse tervise skooris. NHP tulemused olid sarnased, alamkaalal "valu" (vastupidine polaarsus) täheldasime olulist muutust I skaneerimisest II skannimiseks (t (14) a = ?? 5.674, p <0.001, skaneerimine III (t (12 )? =? 7.040, p <0.001 ja skaneerimine IV (t (10)? = ?? 3.258, p? =? 0.009). Samuti leidsime olulise suurenemise alamkaalal "füüsiline liikuvus" alates I skannimisest kuni III skannimiseni (t (12) p = 3.974, p? = 0.002) ja skaneerimine IV (t (10) p = 2.511, p = = 0.031). I ja II skannimise vahel olulist muutust ei olnud ( kuus nädalat pärast operatsiooni).
Struktuurilised andmed
Ristlõike analüüs. Lisasime vanuse kovariaadina üldisesse lineaarsesse mudelisse ja ei leidnud, et vanus segaks. Võrreldes soo ja vanusega sobitatud kontrollidega näitasid puusa primaarse OA-ga patsiendid (n? =? 20) operatsioonieelselt (Scan I) vähenenud halli aine eesmises tsingulaarkoores (ACC), saarekoores, operkumis, dorsolateraalses prefrontaalses ajukoores ( DLPFC), parem ajaline poolus ja väikeaju (tabel 1 ja joonis 1). Välja arvatud parempoolsed putamenid (xa =? 31, y? = ?? 14, zp = ?? 1; p <0.001, tp =? 3.32), ei leitud OA-ga patsientidel halli aine tiheduse olulist suurenemist tervislikele kontrollidele. Võrreldes IV aja skaneerimise patsiente sobitatud kontrollidega, leiti I skaneeringut kasutades ristlõikeanalüüsis kontrollidega võrreldavad tulemused.
Joonis 1: statistilised parameetrilised kaardid, mis demonstreerivad halli aine struktuurseid erinevusi kroonilise valu puhul, mis on tingitud primaarse puusa OA-st võrreldes kontrolliga ja pikema aja jooksul nendega võrreldes. Olulised halli aine muutused on näidatud värvidega, ristlõike andmed on kujutatud punasega ja pikisuunalised andmed kollasega. Telgtasand: pildi vasak pool on aju vasak pool. üles: Puusa primaarse OA-st tingitud kroonilise valuga patsientide ja kontrollimata katsealuste vahel halli aine olulise vähenemise piirkonnad. p <0.001 korrigeerimata põhi: halli aine suurenemine 20 valuvabal patsiendil kolmandal ja neljandal skaneerimisperioodil pärast täielikku puusaliigese asendamise operatsiooni, võrreldes esimese (operatsioonieelse) ja teise (6 nädalat pärast operatsiooni) skaneerimisega. p <8 korrigeerimata graafikud: kontrastsuse hinnangud ja 0.001% usaldusvahemik, huvipakkuvad mõjud, suvalised ühikud. x-telg: kontrastid 90 ajapunkti jaoks, y-telg: kontrastsuse hinnang ACC korral 4, 3, 50 ja insula puhul kontrastsuse hinnang 2, 36, 39.
Vasaku puusa OA-ga patsientide andmete nippimine (n? =? 7) ja nende võrdlemine tervete kontrollidega ei muutnud tulemusi oluliselt, kuid taalamuse vähenemise osas (x? =? 10, y? = ?? 20, z = = 3, p <0.001, t = 3.44) ja parempoolse väikeaju suurenemine (x = = 25, y = 37, z = 50, p <0.001, t =? 5.12), mis ei saavutanud olulisust patsientide kokkuvõtmata andmetes võrreldes kontrollidega.
Pikisuunaline analüüs. Pikisuunalises analüüsis tuvastati halli aine märkimisväärne suurenemine (p <.001 parandamata), võrreldes esimest ja teist skannimist (krooniline valu / operatsioonijärgne valu) kolmanda ja neljanda skaneerimisega (valuvaba) ACC-s, ajukoor, väikeaju ja pars orbitalis OA-ga patsientidel (tabel 2 ja joonis 1). Hallaine vähenes aja jooksul (p <.001 kogu aju analüüs korrigeerimata) OA-ga patsientidel sekundaarses somatosensoorses ajukoores, hipokampuses, keskkõrvalises ajukoores, taalamuses ja kaudaalses tuumas (joonis 2).
Joonis 2: a) Aju halli aine märkimisväärne suurenemine pärast edukat operatsiooni. Puusa primaarse OA-st tingitud kroonilise valuga patsientide halli aine olulise vähenemise telgvaade võrreldes kontrollisikutega. p <0.001 korrigeerimata (ristlõike analüüs), b) Halli aine pikisuunaline suurenemine aja jooksul kollases värvitoonis, võrreldes I ja II skaneerimine III> skaneerimine IV) OA-ga patsientidel. p <0.001 parandamata (pikianalüüs). Pildi vasak pool on aju vasak pool.
Vasaku puusa OA-ga patsientide andmete nippimine (n? =? 7) ei muutnud tulemusi oluliselt, kuid aju halli aine vähenemise osas Heschli Gyrus'is (x? = ?? 41, y? = ?? 21, zp =? 10, p <0.001, tp =? 3.69) ja Precuneus (xa =? 15, yp = ?? 36, zp =? 3, p <0.001, tp =? 4.60). .
Esimese skaneerimise (eeloperatsioon) vastandamisel skaneeringutele 3 + 4 (operatsioonijärgne operatsioon) leidsime, et frontaalse ja motoorse ajukoores on suurenenud halli aine (p <0.001 korrigeerimata). Märgime, et see kontrastsus on vähem range, kuna meil on nüüd vähem skaneeringuid ühe seisundi kohta (valu vs mittevalu). Künnist langetades kordame seda, mida oleme leidnud, kasutades kontrasti 1 + 2 vs 3 + 4.
Otsides piirkondi, mis suurenevad kõigi ajaintervallide jooksul, leidsime koksartroosiga patsientidel aju halli aine muutused motoorsetes piirkondades (piirkond 6) pärast puusa täielikku asendamist (I skaneerimine)dbm.neuro.uni-jena.de/vbm/) võiksime seda leidu korrata eesmises ja keskmise tsingulaadiga ajukoores ning mõlemas esiosas.
Arvutasime efekti suurused ja ristlõikeanalüüs (patsiendid vs kontrollid) andis Coheni SD 1.78751 ACC piigi vokslis (x = = 12, y = = 25, z? = ?? 16). Samuti arvutasime pikisuunalise analüüsi jaoks (kontrastse skaneerimise 1 + 2 vs skaneerimise 3 + 4) Coheni sd. Selle tulemusel saadi ACC-s Coheni SD 1.1158 (x = = 3, y = = 50, z = = 2). Insulaadi (x? = ?? 33, y? =? 21, z? =? 13) ja sama kontrastiga seoses on Coheni sd 1.0949. Lisaks arvutasime ROI-s Coheni sd-kaardi nullist erineva vokseliväärtuste keskmise (mis koosneb Hingardi-Oxfordi kortikaalsest struktuuriatlasest tuletatud tsingulaadi gyrus ja subkallosaalse korteksi esiosast): 1.251223.
Dr Alex Jimenez Insight
Kroonilise valu patsiendid võivad aja jooksul kogeda mitmesuguseid terviseprobleeme, peale nende juba nõrkade sümptomite tekkimise. Näiteks on paljudel inimestel nende valu tõttu magamishäired, kuid mis kõige tähtsam, krooniline valu võib põhjustada ka erinevaid vaimse tervisega seotud probleeme, sealhulgas ärevust ja depressiooni. Valu mõju ajule võib tunduda liiga suur, kuid kasvavad tõendid näitavad, et need aju muutused ei ole püsivad ja neid saab pöörata, kui kroonilised valu patsiendid saavad nende tervisega seotud probleemide nõuetekohast ravi. Artikli järgi ei peegelda kroonilise valu korral leitud hallinähtuse kõrvalekalded ajukahjustust, vaid pigem on need pöörduvad tagajärjed, mis normaliseeruvad, kui valu on adekvaatselt ravitud. Õnneks on saadaval mitmesugused raviviisid, mis aitavad krooniliste valu sümptomite leevendamiseks ja aju struktuuri ja funktsiooni taastamiseks.
Arutelu
Kogu aju struktuuri aja jooksul jälgides kinnitame ja laiendame hiljuti avaldatud pilootandmeid [17]. Leidsime kroonilise valu seisundis primaarse puusaliigese osteoartriidiga patsientidel aju halli aine muutusi, mis pöörduvad osaliselt tagasi, kui neil patsientidel on valu vaba, pärast puusaliigese endoproteesimist. Halli aine osaline suurenemine pärast operatsiooni on peaaegu samades piirkondades, kus enne operatsiooni on täheldatud halli aine vähenemist. Vasaku puusaliigese OA-ga patsientide andmete ümberlükkamine (ja seetõttu valu küljele normaliseerumine) mõjutas tulemusi vähe, kuid lisaks näitas Heschli gyrus ja Precuneus halli aine vähenemist, mida me ei saa lihtsalt seletada kuna a priori hüpoteesi pole olemas, suhtuge sellesse väga ettevaatlikult. Kuid skannimisel I täheldatud erinevus patsientide ja tervete kontrollide vahel oli IV skaneeringu ristlõikeanalüüsis siiski täheldatav. Halli aine suhteline suurenemine aja jooksul on seetõttu peen, st pole piisavalt eristatav, et avaldada mõju ristlõike analüüsile, mis on juba näidatud kogemustest sõltuva plastilisuse uurimise uuringutes [30], [31]. Märgime, et asjaolu, et kroonilise valu põhjustatud aju muutuste osad on pöörduvad, ei välista, et nende muutuste mõned muud osad on pöördumatud.
Huvitaval kombel täheldasime, et kroonilistel valu patsientidel enne operatsiooni kroonilise valu ACC-i hallakese väheneb, tundub, et see kestab 6 nädalat pärast operatsiooni (skannimine II) ja suureneb ainult skaneerides III ja IV, mis võib olla tingitud operatsioonijärgsest valu või motoorika vähenemisest funktsioon. See on kooskõlas NHP-s sisalduva füüsilise liikuvuse skoori käitumisandmetega, mis ei mõjutanud postoperatiivselt II ajahetkel olulisi muutusi, kuid oluliselt suurenesid III ja IV skaneerimise suhtes. Pange tähele, et meie patsiendid ei teatanud pärast operatsiooni puusaliigest valu, kuid kogenud operatsioonijärgset valu ümbritsevatel lihastel ja nahal, mida patsiendid väga erinevalt tundsid. Siiski, kui patsiendid teatasid ikkagi mõningast skaalat skaneeriva II korral, kontrasteeriti ka skaneeringu esialgse skriinimisega (kirurgiline operatsioon) skooriga III + IV (operatsioonijärgne operatsioon), mis näitas halli aine suurenemist esiosa ajukoores ja motoorne ajukoores. Pange tähele, et see kontrastsus on vähem range, kuna skaneerib vähem tingimusi (valu ja mitte valu). Kui me alandasime künnist, kordame seda, mida oleme leidnud I + II ja III + IV kontrasti abil.
Meie andmed viitavad tungivalt sellele, et kroonilise valuga patsientide halli aine muutused, mida tavaliselt leidub supraspinaalse notsitseptiivse töötlemisega seotud piirkondades [4], ei ole tingitud neuronite atroofiast ega ajukahjustustest. Asjaolu, et need kroonilise valu seisundis täheldatud muutused ei pöördu täielikult, võib seletada suhteliselt lühikese vaatlusperioodiga (aasta pärast operatsiooni versus seitse aastat kestnud kroonilist valu enne operatsiooni). Neuroplastilised aju muutused, mis võivad olla välja kujunenud mitme aasta jooksul (pideva notsitseptiivse sisendi tagajärjel), vajavad tõenäoliselt rohkem aega, et täielikult pöörduda. Teine võimalus, miks halli aine suurenemist saab tuvastada ainult pikisuunalistes andmetes, kuid mitte ristlõike andmetes (st kohordide vahel IV ajahetkel), on see, et patsientide arv (n? =? 20) on liiga väike. Tuleb märkida, et mitme indiviidi aju erinevus on üsna suur ja pikisuunaliste andmete eeliseks on see, et dispersioon on suhteliselt väike, kuna samu ajusid skaneeritakse mitu korda. Järelikult on peenmuutused tuvastatavad ainult pikisuunalistes andmetes [30], [31], [32]. Muidugi ei saa me välistada, et need muutused on vähemalt osaliselt pöördumatud, kuigi see on ebatõenäoline, arvestades harjutuste spetsiifilise struktuurilise plastilisuse ja ümberkorraldamise tulemusi [4], [12], [30], [33], [34]. Sellele küsimusele vastamiseks peavad tulevased uuringud uurima patsiente korduvalt pikema aja jooksul, võib-olla ka aastate jooksul.
Pange tähele, et me võime teha piiratud järeldusi morfoloogiliste aju muutuste dünaamika kohta aja jooksul. Põhjuseks on see, et kui me kavandasime seda uuringut 2007-is ja skannimeerisime 2008-is ja 2009-is, ei olnud teada, kas üldiselt toimuvad struktuurimuutused ja teostatavuse huvides valisime siin kirjeldatud skaneerimiskuupäevad ja -ajakavad. Võib väita, et halli aine muutub ajas, mida me kirjeldame ka patsiendirühmale, võib juhtuda ka kontrollrühmas (aja efekt). Kuid kõik muutused vananemise tõttu, kui üldse, peaksid eeldatavasti olema mahu vähenemine. Arvestades meie a priori hüpoteesi, mis põhineb 9i iseseisvatel uuringutel ja kohordidel, kus kroonilise valu patsientidel on halli aineid vähenenud [7], [8], [9], [15], [24], [25], [26], [27], [28] keskendusime me aja jooksul piirkondlikele tõusudele ja usume, et meie leidmine ei ole lihtne ajaline efekt. Pidage meeles, et me ei saa välistada, et aja jooksul vähenes halli mass, mida meie patsiendigrupis leidsime, võib olla ajalise mõju tõttu, sest me ei ole skriininud meie kontrollrühma samal ajajärgul. Tulemused peaksid tulevaste uuringute eesmärgiks olema rohkem ja lühemad ajavahemikud, arvestades, et füüsilise koormusega seotud morfomeetrilised aju muutused võivad ilmneda nii kiiresti kui pärast 1i nädalat [32], [33].
Peale ajukella [17], [34] valu kõrvalnähtude mõju avaldasime, et muutused motoorses funktsioonis mõjutavad tõenäoliselt ka struktuurimuutusi. Me leidsime, et mootori- ja primootorid (ala 6) suurenevad kõigi ajavahemike järel (joonis 3). Intuitiivselt võib see olla tingitud motoorse funktsiooni paranemisest aja jooksul, kuna patsiendid ei olnud tavapärase eluea jooksul enam piiratud. Eelkõige me ei keskendunud motoorsele funktsioonile, vaid valu tekkimise paranemisele, arvestades meie esialgset püüdlust uurida, kas kroonilise valu patsiendil tuntud peaahelas hallatava aine reduktsioon on põhimõtteliselt pöörduv. Seetõttu ei kasutanud me spetsiaalseid vahendeid motoorse funktsiooni uurimiseks. Siiski on valusündroomide all kannatavate patsientide (funktsionaalne) motoorika reorganiseerimine hästi dokumenteeritud [35], [36], [37], [38]. Peale selle on motoorset ajukoekstrakti üheks eesmärgiks terapeutilisel lähenemisel meditsiiniliselt raskesti ligipääsetavate kroonilise valuga patsientidel, kes kasutavad otsest ajude stimulatsiooni [39], [40], transkraniaalset otsesisene voolu stimulatsiooni [41] ja korduvat transkraniaalset magnetilist stimulatsiooni [42], [43]. Sellise modulatsiooni täpne mehhanism (soodustamine vs inhibeerimine või lihtsalt häireid valuga seotud võrkudes) pole veel välja selgitatud [40]. Hiljutine uuring näitas, et konkreetne motoorika kogemus võib muuta aju struktuuri [13]. Sünoptogenees, liikumisharjumuste ümberkorraldamine ja angiogenees motoorses koorega võib esineda motoorse ülesande erinõuetega. Tsao et al. näitasid kroonilise alaseljavalu all kannatavate patsientide motoorses koorega ümberkorraldamist, mis näivad olevat seljavalu-spetsiifilised [44] ja Puri et al. täheldati vasaku täiendava mootorikoha halli ainevahetuse vähenemist fibromüalgia põdejatele [45]. Meie uuring ei olnud mõeldud selleks, et lahti seletada erinevaid tegureid, mis võivad aju muutuda kroonilises valu, kuid me tõlgendame oma andmeid halli aine muutuste kohta, et need ei peegelda ainult konstantse notsitseptiivse sisendi tagajärgi. Hiljutine uuring neuropaatilise valuga patsientidel tõi esile ebanormaalsed ajuspiirkonnad, mis hõlmavad emotsionaalset, autonoomset ja valulist tajumist, mis tähendab, et neil on kroonilise valu [28] ülemaailmses kliinilises pildis kriitiline roll.
Joonis 3: Statistilised parameetrikaardid, mis näitavad aju halli aine olulist suurenemist motoorsetes piirkondades (6. piirkond) koksartroosiga patsientidel enne THR-i järgset (pikianalüüs, skann I Kontrastsuse hinnangul väärtus x x = = 19, y = = 12, z = = 70.
Kaks hiljutist pilootuuringut keskendusid osteoartriidi patsientidele puusa-asendusravi, ainus kroonilise valusündroomi, mis on põhiliselt puusaliigese [17] ja [46] ravitavaks ning nende andmetega kaasneb hiljuti kroonilise alaseljavalu patsientide uuring [ 47]. Neid uuringuid tuleb vaadelda mitmete pikisuunaliste uuringute valguses, milles uuritakse inimese kogemata sõltuvat neuronaalset plastilisust [30], [31] ja hiljutist uuringut strukturaalsete aju muutuste kohta tervetel vabatahtlikel, kellel esineb korduv valulik stimulatsioon [34] . Kõigi nende uuringute peamine sõnum on see, et valu valu ja patsiendi kontrollide vaheline aju struktuuri peamine erinevus võib paranemise tagajärjel taanduda. Siiski tuleb arvestada, et lihtsalt ei ole selge, kas kroonilise valuga patsientide muutused on tingitud üksnes notsitseptiivsest sisendist või valu tagajärgedest või mõlemast. Aju [6], [12], [28], [48]) kujundamiseks on enam kui tõenäoline, et käitumisharjumused, nagu näiteks sotsiaalsete kontaktide puudumine või paranemine, agility, füüsiline väljaõpe ja elustiili muutused, on piisavad. Eriti on depressioon kaasne haigestumus või valu tagajärg, et selgitada erinevusi patsientide ja kontrollide vahel. Väikesel rühmal meie OA-ga patsientidest ilmnesid kerge kuni mõõdukas depressiivsed sümptomid, mis aja jooksul muutuvad. Me ei leidnud BDI-skoori kohta märkimisväärseid struktuurseid muutusi, kuid tekib küsimus, kui palju muid käitumisharjumusi valu ja motoorika paranemise tõttu võivad kaasa aidata tulemustele ja millises ulatuses nad seda teevad. Need käitumisharjumused võivad mõjutada kroonilise valu halva aine vähenemist ja halli aine suurenemist, kui valu on kadunud.
Teine oluline tegur, mis võib tulemuste tõlgendamist erineda, on asjaolu, et peaaegu kõik kroonilise valu patsiendid võtsid ravimeid valu vastu, mille nad lõpetasid, kui nad olid valu vaba. Võib väita, et mittesteroidsed põletikuvastased ravimid, nagu diklofenak või ibuprofeen, mõjutavad närvisüsteeme mõnevõrra, ja see kehtib ka opioidide, epilepsiavastaste ravimite ja antidepressantide kohta, mida kasutatakse sageli kroonilise valu korral. Mõõdukate ja muude ravimite mõju morfomeetrilistele leidetele võib olla tähtis (48). Ükski uuring ei ole seni näidanud valu ravimite mõjusid aju morfoloogiale, kuid paljudes dokumentides leiti, et kroonilise valu patsientidel aju struktuuri muutusi ei seostata mitte ainult valu-seotud mitteaktiivsusega [15] ega valu ravimitega [7], [9], [49]. Siiski puuduvad eriuuringud. Edasised uuringud peaksid keskenduma kortikaalse plastilisusega seotud kogemustest sõltuvatele muutustele, millel võib olla kroonilise valu raviks ulatuslik kliiniline tähendus.
Me leidsime ka pikaajalisel analüüsil halli aine hulga vähenemise, mis võib olla tingitud reorganiseerimisprotsessidest, mis kaasnevad muutustega motoorsete funktsioonide ja valu tajumisel. Aju halli aine pikisuunaliste muutuste kohta valuvormides on vähe teavet, mistõttu pole meil hüpoteesi nende alade halli aeglase vähenemise pärast operatsiooni. Teutsch et al. [25] leidis tervetel vabatahtlikel, et täisprotokollis oli kaheksa järjestikuse päeva jooksul valus stimulatsioon ajuhaiguse suurenemist somatosensory ja midcingual cortex. Halli aine sisalduse suurenemine pärast eksperimentaalset noiteseptiivset sisendit kattis anatoomiliselt mingil määral ajuhaige aine vähenemisega selles uuringus patsientidel, keda oli ravitud pikaajalisest kroonilisest valu. See tähendab, et tervetele vabatahtlikele nakatamatu sisendiga kaasneb füüsilise koormusega seotud strukturaalsed muutused, kuna see võib olla kroonilise valuga patsientidel ja et need muutused pöörduksid tervetel vabatahtlikel, kui natsitseptiivne sisestus lõpeb. Sellest tulenevalt võib OA-ga patsientide halli ainete vähenemist neis piirkondades järgida sama põhiprotsessiga: aju muutused sõltuvad füüsilisest koormusest [50]. Mitte-invasiivse protseduurina on MR Morphometry ideaalne vahend haiguste morfoloogiliste substraatide leidmiseks, süvendades meie arusaamist aju struktuuri ja funktsiooni vahelistest suhetest ja isegi terapeutiliste sekkumiste jälgimiseks. Üks tuleviku suurtest väljakutsetest on kohandada seda võimsat vahendit kroonilise valu multitsentrilisteks ja terapeutilisteks uuringuteks.
Selle uuringu piirangud
Kuigi see uuring on meie eelmise uuringu laiendus, laiendades jälgimisandmeid 12 kuuni ja uurides rohkem patsiente, on meie põhimõte, et kroonilise valu morfomeetrilised aju muutused on pöörduvad, üsna peen. Mõju suurused on väikesed (vt eespool) ja mõjud on osaliselt ajendatud piirkondliku aju halli aine mahu vähenemisest skaneerimise ajahetkel 2. Kui me jätame skaneerimisest 2 andmed välja (vahetult pärast operatsiooni), on oluline aju halli aine suurenemine motoorses ja frontaalses ajukoores elab üle künnise p <0.001 korrigeerimata (tabel 3).
Järeldus
Ei ole võimalik eristada, mil määral täheldatud struktuursed muutused on tingitud noiteseptiivse sisendi muutustest, motoorse funktsiooni muutustest või ravimi tarbimisest või heaolu muutumisest kui sellisest. Esimese ja viimase skaneerimisega grupi kontrastide varjamine üksteisega näitas tunduvalt vähem oodatavaid erinevusi. Võimalik, et aju muutused kroonilise valu ja kõigi tagajärgede tõttu tekivad üsna pikka aja jooksul ja vajavad ka tagasi pöördumist. Sellegipoolest näitavad need tulemused ümberkorraldamisprotsessid, mis näitavad kindlalt, et krooniline noiteseptiivne sisend ja motoorika kahjustus neis patsientidel toovad kortikaalsetes piirkondades muutunud töötlemise ja järelikult ka aju muutused, mis on põhimõtteliselt pööratavad.
Tunnustused
Täname kõiki vabatahtlikke selles uuringus osalemise eest ja Hamburgi NeuroImage Nord füüsika ja meetodite rühma. Uuringule anti eetiline nõusolek kohaliku eetikakomitee poolt ja enne uurimist saadi kirjalik teadlik nõusolek kõigist uuringus osalenutest.
Rahastamise aruanne
Seda tööd toetasid DFG (Saksamaa Teadusfond) (MA 1862 / 2-3) ja BMBF (Saksamaa Liitvabariigi Haridus- ja Teadusministeerium) toetused (371 57 01 ja NeuroImage Nord) toetused. Rahastajatel ei olnud rolli uuringute kujundamisel, andmete kogumisel ja analüüsimisel, käsutusse andmise otsuse tegemisel ega käsikirjali koostamisel.
Endokannabinoidne süsteem: hädavajalik süsteem, millest te pole kunagi kuulnud
Juhul, kui te ei ole endocannabinoid-süsteemi või ECS-i kuulnud, pole vaja piinlikkust tunda. 1960-i tagasi jätsid uurijad, kes olid huvitatud kanepi bioaktiivsusest, lõpuks isoleerima paljusid aktiivseid kemikaale. Kuid loomade mudelite uurimiseks uurijatel, kes otsisid selliste ECS-i kemikaalide retseptorit näriliste ajus, leidis see veel kord 30 aastat, mis avas kogu ECS-i retseptorite olemasolu uurimise maailma ja selle, milline on nende füsioloogiline eesmärk.
Nüüd teame, et enamikul loomadel, alates kalast kuni lindudeni imetajatele, on endokannabininoid ja me teame, et inimesed ei tooda mitte ainult oma kanabinoide, mis suudavad seda konkreetset süsteemi siduda, vaid toodame ka teisi ühendeid, mis toimivad ECSiga, mida täheldatakse paljude erinevate taimede ja toitude puhul, mis ulatuvad kaugemale kui kanepiliigid.
Inimese keha süsteemina ei ole ECS isoleeritud struktuuriline platvorm nagu närvisüsteem või kardiovaskulaarne süsteem. Selle asemel on ECS retseptorite kogum, mis laialdaselt levib kogu kehas, mis aktiveeritakse ligandide komplekti kaudu, mida me kollektiivselt teame endokannabinoidide või endogeensete kannabinoidide hulgast. Mõlemat kontrollitud retseptorit kutsutakse lihtsalt CB1 ja CB2, kuigi on olemas ka muud, mis olid välja pakutud. PPAR ja TRP-kanalid on vahendajad ka mõned funktsioonid. Samamoodi leiate kaht hästi dokumenteeritud endokannabinoidi: anadamiidi ja 2-arahidinoüülglütserooli või 2-AG.
Lisaks sellele on endokanabinoidide süsteemis põhiliselt ensüümid, mis sünteesivad ja murravad endokannabinoide. Arvatakse, et endokannabinoide sünteesitakse vajalikus vundamendis. Esmaste ensüümide hulka kuuluvad diatsüülglütserool-lipaas ja N-atsüülfosfatidüületanoolamiin-fosfolipaas D, mis vastavalt sünteesivad 2-AG ja anandamiidi. Kaks peamist degradeerivat ensüümi on rasvhappe amiidi hüdrolaas või FAAH, mis lagundab anandamiidi ja monoatsüülglütserooli lipaasi või MAGL-i, mis lagundab 2-AG. Nende kahe ensüümi reguleerimine võib suurendada või vähendada ECS modulatsiooni.
Mis on ECSi funktsioon?
ECS on keha peamine homeostaatiline reguleerimissüsteem. Seda võib kergesti vaadelda kui keha sisemist adaptogeenset süsteemi, mis alati töötab erinevate funktsioonide tasakaalu säilitamiseks. Endokannabinoidid toimivad üldjuhul neuromodulaatorina ja reguleerivad enamasti paljusid kehasisesi protsesse, alates viljastumisest kuni valu. Mõned neist ECS-i tuntumatest funktsioonidest on järgmised:
Närvisüsteem
Kesknärvisüsteemist või kesknärvisüsteemist inhibeerib CB1 retseptorite üldine stimulatsioon glutamaadi ja GABA vabanemist. Kesknärvisüsteemil mängib ECS mälu moodustamisel ja õppimisel rolli, edendab neurogeneesi hippokampuses, reguleerib samuti neuronaalset ärrituvust. ECS osaleb ka selles, kuidas aju reageerib vigastustele ja põletikele. Alates seljaaju modifitseerib ECS valusignaali ja suurendab looduslikku analgeesiat. Perifeerses närvisüsteemis, milles CB2 retseptorid kontrollivad, toimib ECS peamiselt sümpaatilise närvisüsteemis, et reguleerida soole-, kuse- ja reproduktiivrakkude funktsioone.
Stress ja meeleolu
ECS-il on mitu mõju stressireaktsioonidele ja emotsionaalsele reguleerimisele, näiteks selle kehalise reageerimise algatamine akuutsele stressile ja aja jooksul kohanemine pikemaajaliste emotsioonidega, nagu hirm ja ärevus. Tervislik endokannabinoidsüsteem on kriitiline selle üle, kuidas inimesed moduleerivad rahuldava ärritusastme taset võrreldes ülemäärase ja ebameeldiva tasemega. ECS mängib ka rolli mälu moodustamisel ja võib-olla eriti selle poolest, kuidas aju jäljendab stressi või vigastusi mälestustest. Kuna ECS moduleerib dopamiini, noradrenaliini, serotoniini ja kortisooli vabanemist, võib see mõjutada ka emotsionaalset reaktsiooni ja käitumist.
Seedeelundkond
Seedetrakt on varustatud nii CB1i kui ka CB2i retseptoritega, mis reguleerivad geograafiliste tähiste tervise mitmeid olulisi aspekte. Arvatakse, et ECS võib olla "puuduv lüli" seedetrakti funktsionaalset tervist mängivat soolestiku aju-immuunsüsteemi seost kirjeldades. ECS on soolestiku immuunsuse regulaator, võib-olla piirates immuunsüsteemi tervisliku taimestiku hävitamise ja tsütokiinide signalisatsiooni modulatsiooni kaudu. ECS moduleerib looduslikku põletikulist vastust seedetraktil, millel on oluline mõju paljudele terviseküsimustele. Mao-ja üldist geeni-liikumisvõimet näib ka osaliselt reguleeritud ECS.
Söögiisu ja ainevahetus
ECS, eriti CB1 retseptorid, osaleb isu, ainevahetuse ja keharasva reguleerimises. CB1i retseptorite stimuleerimine tõstab toidust otsivat käitumist, suurendab tundlikkust lõhna suhtes, reguleerib ka energia tasakaalu. Mõlemal loomal ja ülekaalulisel inimesel on ECS-i düsregulatsioon, mis võib viia selle süsteemi hüperaktiivsusele, mis aitab kaasa ületamisele ja vähendab energiakulusid. On näidatud, et anandamiidi ja 2-AG tsirkuleerivad tasemed on rasvumisega seotud, mis võib olla osaliselt tingitud FAAH lagundava ensüümi vähenenud tootmisest.
Immuunhaigused ja põletikuline reaktsioon
Immuunsüsteemi rakud ja organid on rikas endokannabinoidi retseptoritega. Kannabinoidi retseptorid on ekspresseeritud vöötohatis, põrnas, mandlites ja luuüdis, samuti T- ja B-lümfotsüütidel, makrofaagidel, nuumrakkudel, neutrofiilidel ja loomulikes tapjarakkudes. ECS-i peetakse immuunsüsteemi tasakaalu ja homöostaasi esmaseks juhiks. Kuigi kõiki ECS-i funktsioone immuunsüsteemist ei mõisteta, tundub, et ECS reguleerib tsütokiinide tootmist ja mängib rolli immuunsüsteemi ülekaalulisuse ennetamisel. Põletik on immuunvastuse loomulik osa ja see mängib väga tavalist rolli kehas ägedate solvangute, sealhulgas vigastuste ja haiguste puhul; sellegipoolest, kui seda ei kontrollita, võib see muutuda krooniliseks ja aidata kaasa tervisehäirete, nagu krooniline valu, kaskaadile. Kontrollides immuunvastust, aitab ECS säilitada organismi tasakaalustatumat põletikuvastast toimet.
Muud ECSi reguleeritud tervishoiuvaldkonnad:
Luu tervis
Viljakus
naha tervise
Arteriaalne ja hingamisteede tervis
Uni ja ööpäevane rütm
Tervisliku ECS-i parimaks toetamiseks on küsimus, mida paljud teadlased üritavad nüüd vastata. Hoidke lisateavet selle tekkiva teema kohta.
KokkuvõtteksKroonilist valu on seostatud aju muutustega, sealhulgas halli aine vähenemisega. Kuid ülaltoodud artikkel näitas, et krooniline valu võib muuta aju üldist struktuuri ja funktsiooni. Kuigi krooniline valu võib nendeni viia, võib muude terviseprobleemide kõrval patsiendi põhisümptomite õige ravi aju muutused tagasi pöörata ja halli ainet reguleerida. Pealegi on endokannabinoidsüsteemi tähtsuse taga olnud üha rohkem uurimusi ja see toimib nii kroonilise valu kui ka muude terviseprobleemide kontrollimisel ja haldamisel. Riikliku biotehnoloogia teabe keskuse (NCBI) viidatud teave. Meie teabe ulatus piirdub kiropraktika, samuti selgroovigastuste ja seisunditega. Teema arutamiseks küsige palun dr Jimenezilt või võtke meiega ühendust aadressil915-850-0900.
Juhib dr Alex Jimenez
Täiendavad teemad: Seljavalu
Seljavalu on üks kõige levinumaid puudeid ja töölt puuduvaid tööpäevi kogu maailmas. Tegelikkuses on seljavalu olnud ka arstikeskuse külastuste teine kõige levinum põhjus, millest arvukus oli ainult ülemiste hingamisteede nakkustega. Ligikaudu 80 protsenti elanikkonnast kogeb teatud tüüpi seljavalu vähemalt korra kogu eluea jooksul. Lülisamba on keeruline struktuur, mis koosneb teistest pehmetest kudedest koosnevate luude, liigeste, sidemete ja lihaste vahel. Selle tagajärjel tekivad vigastused ja / või raskendavad seisundid, näiteks hernide kettad, võib lõpuks põhjustada seljavalu sümptomeid. Sageli on seljavalu kõige sagedasem spordivigastus või autoõnnetusjuhtumite põhjus, kuid mõnikord võib kõige lihtsam liikumine olla valulike tulemustega. Õnneks võivad alternatiivsed ravivõimalused, nagu kiropraktiline ravi, aidata leevendada seljavalu, kasutades selleks spinaalset reguleerimist ja manuaalseid manipulatsioone, parandades lõpuks valu hõlbustamist.
3. Wrigley PJ, Gustin SM, Macey PM, Nash PG, Gandevia SC jt. (2009) Inimese motoorika ja motooriaalsete radade anatoomilised muutused täielikus rindkere stenokardia vigastusest.Cereb Cortex19: 224 232. [PubMed]
4. Mai A (2008) Krooniline valu võib muuta aju struktuuri.Valu137: 7 15. [PubMed]
5. Mai A (2009) Morfing vokselid: peavalu patsientide strukturaalse kuvamise peapööritus. Aju[PubMed]
6. Apkarian AV, Baliki MN, Geha PY (2009)Kroonilise valu teooria suunas.Prog Neurobiol87: 81 97. [PMC tasuta artikkel][PubMed]
7. Apkarian AV, Sosa Y, Sonty S, Levy RM, Harden RN jt. (2004)Krooniline seljavalu on seotud vähenenud prefrontaalse ja talamiinse halli aine tihedusega.J Neurosci24: 10410 10415. [PubMed]
8. Rocca MA, Ceccarelli A, Falini A, Colombo B, Tortorella P jt. (2006)Aju-halli aine muutused migreenihaigetel, kellel on T2-i nähtavad kahjustused: 3-T MRI uuring.Insult37: 1765 1770. [PubMed]
9. Kuchinad A, Schweinhardt P, Seminowicz DA, Wood PB, Chizh BA jt. (2007)Kiired ajuhaiguse kadu fibromüalgiaga patsientidel: aju enneaegne vananemine?J Neurosci27: 4004-4007.[PubMed]
10. Tracey I, Bushnell MC (2009)Kuidas neuroimaging uuringud on meile proovinud ümber mõelda: kas krooniline valu on haigus?J Pain10: 1113 1120. [PubMed]
11. Franke K, Ziegler G, Kloppel S, Gaser C (2010)T1-kaalutud MRI-skannimisega, kasutades kerneli meetodeid, tervete isikute vanuse hindamine: erinevate parameetrite mõju uurimine.Neuroimage50: 883 892. [PubMed]
12. Draganski B, mai A (2008)Koolitusest tingitud struktuursed muutused täiskasvanud inimese ajus.Behav Brain Res192: 137 142. [PubMed]
13. Adkins DL, Boychuk J, Remple MS, Kleim JA (2006)Mootorikoolitus tekitab moodsa koore ja seljaaju vahelist kogemust iseloomustavat plastilisust.J Appl Physiol101: 1776 1782. [PubMed]
14. Duerden EG, Laverdure-Dupont D (2008)Praktika muudab ajukooreks.J Neurosci28: 8655 8657. [PubMed]
15. Draganski B, Moser T, Lummel N, Ganssbauer S, Bogdahn U jt. (2006)Talamiinhalli massi vähenemine pärast jäseme amputatsiooni.Neuroimage31: 951 957. [PubMed]
16. Nikolajsen L, Brandsborg B, Lucht U, Jensen TS, Kehlet H (2006)Kogu puusaliigese kopsuarteri põhjustatud krooniline valu: üleriigiline küsimustik.Acta Anestesiol Scand50: 495 500. [PubMed]
17. Rodriguez-Raecke R, Niemeier A, Ihle K, Ruether W, mai A (2009)Aju-halli aine kroonilise valu langus on pigem tagajärg, mitte valu põhjus.J Neurosci29: 13746 13750. [PubMed]
18. Beck AT, Ward CH, Mendelson M, Mock J, Erbaugh J (1961)Inventar depressiooni mõõtmiseks.Arch Gen Psychiatry4: 561 571. [PubMed]
19. Franke G (2002) Die Symptom-Checkliste nach LR Derogatis - Manual. Göttingen Beltzi test Verlag.
20. Geissner E (1995) Valu tajumise skaala - diferentseeritud ja muutustele tundlik skaala kroonilise ja ägeda valu hindamiseks. Taastusravi (Stuttg) 34: XXXV XLIII. [PubMed]
21. Bullinger M, Kirchberger I (1998) SF-36 - Fragebogen zum Gesundheitszustand. Käsitsi anweisung. Gttingen: Hogrefe.
23. Hea CD, Johnsrude IS, Ashburner J, Henson RN, Friston KJ jt. (2001)Vokselipõhine morfomeetriline uuring vananemise kohta 465i normaalsete täiskasvanud inimese ajus.Neuroimage14: 21 36. [PubMed]
24. Baliki MN, Chialvo DR, Geha PY, Levy RM, Harden RN jt. (2006)Krooniline valu ja emotsionaalne aju: spetsiifiline ajutegevus, mis on seotud spontaansete kroonilise seljavalu intensiivsuse kõikumistega.J Neurosci26: 12165 12173. [PMC tasuta artikkel][PubMed]
25. Lutz J, Jager L, de Quervain D, Krauseneck T, Padberg F jt. (2008)Fibromüalgiat põdevate patsientide aju valgel ja hallmalmil esinevad kõrvalekalded: difusioon-tensori ja ruumilise pildistamise uuring.artriit Rheum58: 3960 3969. [PubMed]
26. Wrigley PJ, Gustin SM, Macey PM, Nash PG, Gandevia SC jt. (2008) Inimese mootorikorteede ja motoorikatte anatoomilised muutused täieliku rindkere omavolilise kahjustuse järel.Cereb Cortex19: 224 232. [PubMed]
27. Schmidt-Wilcke T, Hierlmeier S, Leinisch E (2010) Muutunud piirkondlik aju morfoloogia kroonilise näovaluga patsientidel. Peavalu[PubMed]
28. Geha PY, Baliki MN, Harden RN, Bauer WR, Parrish TB jt. (2008)Aju krooniline CRPS valu: ebanormaalne hall-valge aine vastasmõju emotsionaalsetes ja autonoomsetes piirkondades.Neuron60: 570 581. [PMC tasuta artikkel][PubMed]
29. Brazier J, Roberts J, Deverill M (2002)SF-36i eelistuste alusel põhineva tervisemõõtmise hinnang.J Health Econ21: 271 292. [PubMed]
30. Draganski B, Gaser C, Busch V, Schuierer G, Bogdahn U jt. (2004)Neuroplasticity: muutused halli aine tekitatud koolituse.loodus427: 311 312. [PubMed]
31. Boyke J, Driemeyer J, Gaser C, Buchel C, mai A (2008)Koolitusest tingitud aju struktuur muutub eakatel.J Neurosci28: 7031 7035. [PubMed]
32. Driemeyer J, Boyke J, Gaser C, Buchel C, mai A (2008)Õppimise põhjustatud muutused hallis olekus.PLoS ONE3: e2669. [PMC tasuta artikkel][PubMed]
33. Mai A, Hajak G, Ganssbauer S, Steffens T, Langguth B jt. (2007)Struktuursed ajamuutused pärast 5 päeva sekkumist: dünaamilised aspektid neuroplasticity.Cereb Cortex17: 205 210. [PubMed]
34. Teutsch S, Herken W, Bingel U, Schoell E, mai A (2008)Ajuhaljaga aine muutused korduva ja valuliku stimulatsiooni tõttu.Neuroimage42: 845 849. [PubMed]
36. Flor H, Denke C, Schaefer M, Grusser S (2001)Meelepärase diskrimineerimise koolituse mõju kortikaalsele reorganisatsioonile ja fantoomjäsemevalu.Lantsett357: 1763 1764. [PubMed]
38. Maihofner C, parun R, DeCol R, Binder A, Birklein F jt. (2007)Mootorisüsteem näitab adaptiivseid muutusi keerulises piirkondlikus valusündroomis.Aju130: 2671 2687. [PubMed]
39. Fontaine D, Hamani C, Lozano A (2009)Kroonilise neuropaatilise valu motoorset koore kroonilist stimulatsiooni efektiivsus ja ohutus: kirjanduse kriitiline ülevaade.J Neurosurg110: 251 256. [PubMed]
41. Antal A, Brepohl N, Poreisz C, Boros K, Csifcsak G jt. (2008)Transkraniaalne otsene voolu stimulatsioon somatosensorilise koorega vähendab eksperimentaalselt indutseeritud ägeda valu tajumist.Clin J Pain24: 56 63. [PubMed]
42. Teepker M, Hotzel J, Timmesfeld N, Reis J, Mylius V jt. (2010) Migreeni profülaktilise ravi puhul madala sagedusega rTMS.Cephalalgia30: 137 144. [PubMed]
43. Connell N, Wand B, Marston L, Spencer S, Desouza L (2010)Kroonilise valu mitteinvasiivsed aju stimulatsiooni meetodid. Cochrane'i süstemaatilise ülevaate ja metaanalüüsi aruanne.Eur J Phys Rehabil Med47: 309 326. [PubMed]
44. Tsao H, Galea MP, Hodges PW (2008)Mootorsaagise reorganiseerimine on seotud posturaalse kontrolli puudujääkidega, mis kaasnevad tagasiulatuva alaseljavaluga.Aju131: 2161 2171. [PubMed]
45. Puri BK, Agour M, Gunatilake KD, Fernando KA, Gurusinghe AI jt. (2010) Vasaku täiendava mootorikoha halli ainevahetus täiskasvanud naissoost fibromüalgiaga patsientidel, kellel on märgatav väsimus ja ilma afektiivse häireta: kontrollitud 3-T magnetresonantsuuringute vokselipõhine morfomeetria uuring.J Int Med Res38: 1468 1472. [PubMed]
46. Gwilym SE, Fillipini N, Douaud G, Carr AJ, Tracey I (2010) Puusa valuliku artroosiga seotud taalamuse atroofia on pärast artroplastikat pöörduv; vokselil põhinev pikisuunaline morfomeetriline uuring. Artriit Rheum[PubMed]
47. Seminowicz DA, Wideman TH, Naso L, Hatami-Khoroushahi Z, Fallatah S jt. (2011) Kroonilise alaseljavalu efektiivne ravi inimestel muudab ebanormaalse aju anatoomia ja funktsiooni.J Neurosci31: 7540 7550. [PubMed]
48. Mai A, Gaser C (2006)Magnetresonantsipõhine morfomeetria: aken aju struktuuri plastiliseks.Curr Opin Neurol19: 407 411. [PubMed]
49. Schmidt-Wilcke T, Leinisch E, Straube A, Kampfe N, Draganski B jt. (2005) Kroonilise pinge tüüpi peavaluga patsientidel on halli ainevahetus vähenenud.Neuroloogia65: 1483 1486. [PubMed]
50. Mai A (2009) Morfingu vokselid: peavalu patsientide strukturaalse kuvamise peapööritus.Aju 132 (Pt6): 1419 1425. [PubMed]
Valu biokeemia:Kõigil valusündroomidel on põletikuprofiil. Põletikuline profiil võib inimeselt erineda ja võib erineda ka ühe inimese ajal ja ajal. Valusündroomide ravi on selle põletikuprofiili mõistmine. Valusündroome ravitakse meditsiiniliselt, kirurgiliselt või mõlemat. Eesmärk on pärssida / pärssida põletikuliste vahendajate tootmist. Ja edukas tulemus on see, mille tulemuseks on vähem põletikku ja loomulikult vähem valu.
Valu biokeemia
Eesmärgid:
Kes on peamised mängijad
Millised on biokeemilised mehhanismid?
Millised on tagajärjed?
Põletikuvastane ülevaade:
Key Mängijad
Miks mu õlg valutab? Ülevaade õlavalu neuroanatoomilisest ja biokeemilisest alusest
ABSTRACT
Kui patsient küsib, miks mu õlg valutab?, Pöördub vestlus kiiresti teadusteooria ja mõnikord põhjendamatu oletuse poole. Sageli saab kliinik teada oma selgituste teadusliku aluse piiridest, näidates, et meie arusaam õlavalu olemusest on puudulik. Selles ülevaates kasutatakse süstemaatilist lähenemist, et aidata vastata õlavaluga seotud põhiküsimustele, et anda ülevaade tulevastest uuringutest ja uudsetest meetoditest õlavalu ravimiseks. Uurime (1) perifeersete retseptorite, (2) perifeersete valude töötlemise või "notsitseptsiooni", (3) seljaaju, (4) aju, (5) retseptorite paiknemist õlas ja (6) rolle ) õla närvianatoomia. Samuti kaalume, kuidas need tegurid võivad aidata kaasa õlavalu kliinilise esituse, diagnoosi ja ravi varieeruvusele. Sel moel on meie eesmärk anda ülevaade perifeerse valu tuvastamise süsteemi komponentidest ja kesksest valu töötlemise mehhanismidest õlavalu korral, mis interakteeruvad kliinilise valu tekitamiseks.
SISSEJUHATUS: VÄGA LÜHENDATUD KLIINIKUTE OLULINE VAHETULINE TEADUS
Valu olemus on viimase sajandi jooksul olnud palju poleemikat tekitanud. 17. sajandi Descartesi teoorias1 tehti ettepanek, et valu intensiivsus on otseselt seotud sellega seotud koekahjustuse suurusega ja et valu töödeldakse ühel eraldi rajal. Paljud varasemad teooriad tuginesid sellele nn dualistlikule Descartia filosoofiale, nähes valu ajus toimuva spetsiifilise perifeerse valu retseptori stimuleerimise tagajärjena. 20. sajandil toimus kahe vastandliku teooria, nimelt spetsiifilisuse teooria ja mustriteooria vahel teaduslik lahing. Descartiani spetsiifilisuse teooria nägi valu kui spetsiifilist sensoorse sisendi modaalsust koos oma aparaadiga, samas kui mustri teooria leidis, et valu tuleneb mittespetsiifiliste retseptorite intensiivsest stimulatsioonist. 2. aastal avaldasid Wall ja Melzack 1965 värava valu teooria andis tõendeid mudeli kohta, kus valu tajumist moduleerisid nii sensoorne tagasiside kui ka kesknärvisüsteem. Veel üks tohutu edasiminek valuteoorias avastas samal ajal opioidide toimimisviisi. Seejärel on neurokujutiste ja molekulaarmeditsiini hiljutised edusammud meie üldist arusaama valust oluliselt laiendanud.
Niisiis, kuidas see on seotud õlavalu?Õlavalu on tavaline kliiniline probleemja patsiendi valu kõige paremaks diagnoosimiseks ja raviks on hädavajalik mõista keha valutöötlust. Valutöötlemise alaste teadmiste areng lubab selgitada patoloogia ja valu tajumise ebakõla, see võib aidata meil selgitada ka seda, miks teatud patsiendid teatud ravile ei reageeri.
PÕHJALISED SÜNNITUSKONSTRUKTSIOONID
Perifeersed sensoorsed retseptorid: mehhanoretseptor ja notsitseptor
Inimese lihas-skeleti süsteemis on arvukalt perifeersete sensoorsete retseptorite tüüpe. Neid võib klassifitseerida nende funktsionaalsuse (mehaanoretseptorid, termoretseptorid või notsitseptorid) või morfoloogia (vabad närvilõpmed või erinevat tüüpi kapseldatud retseptorid) põhjal. 5 Erinevaid retseptoritüüpe saab seejärel alamklassifitseerida teatud keemiliste markerite olemasolu. Näiteks retseptorite erinevate funktsionaalsete klasside vahel on märkimisväärseid kattumisi
Perifeerse valu töötlemine: "Notsitsepteerimine"
Kudede kahjustus hõlmab kahjustatud rakkude vabanemist mitmesugustest põletikulistest vahendajatest, sealhulgas bradükiniin, histamiin, 5-hüdroksütrüptamiin, ATP, lämmastikoksiid ja teatud ioonid (K + ja H +). Arahhidoonhappe raja aktiveerimine viib prostaglandiinide, tromboksaanide ja leukotrieenide tootmiseni. Samuti vabanevad tsütokiinid, sealhulgas interleukiinid ja kasvajanekroosifaktor a, ning neurotrofiinid, näiteks närvikasvufaktor (NGF), mis on tihedalt seotud põletiku hõlbustamisega. 15 Muud ained, nagu ergutavad aminohapped (glutamaat) ja opioidid ( endoteliin-1) on seotud ka ägeda põletikulise reaktsiooniga.16 17 Mõned neist toimeainetest võivad otseselt aktiveerida notsitseptoreid, teised aga värbavad teisi rakke, mis vabastavad seejärel täiendavaid soodustavaid aineid.18 See lokaalne protsess põhjustab suurenenud reageerimisvõimet. notsitseptiivsete neuronite normaalse sisendini ja / või vastuse värbamist tavaliselt alammäärale vastavatele sisenditele nimetatakse “perifeerseks sensibiliseerimiseks”. Joonisel 1 on kokku võetud mõned peamised kaasatud mehhanismid.
NGF-l ja transientse retseptori potentsiaalse katioonkanali V-alamperekonna V liikme 1 (TRPV1) retseptoril on sümbiootiline seos põletiku ja notsitseptori sensibiliseerimise osas. Põletikulises koes toodetud tsütokiinid suurendavad NGF tootmist. 19 NGF stimuleerib nuumrakkude poolt histamiini ja serotoniini (5-HT3) vabanemist ning sensibiliseerib ka notsitseptoreid, muutes võib-olla A? kiududest nii, et suurem osa neist saab notsitseptiivseks. TRPV1 retseptor esineb primaarsete aferentsete kiudude alampopulatsioonis ja seda aktiveerivad kapsaitsiin, kuumus ja prootonid. TRPV1 retseptor sünteesitakse aferentse kiu rakukehas ja see transporditakse nii perifeersesse kui ka tsentraalsesse terminali, kus see aitab kaasa notsitseptiivsete aferentide tundlikkusele. Põletiku tagajärjel tekib perifeerselt NGF, mis seondub siis notsitseptori klemmides oleva türosiini kinaasi retseptori 1. tüüpi retseptoriga, seejärel transporditakse NGF rakukehasse, kus see viib TRPV1 transkriptsiooni ülesreguleerimiseni ja sellest tulenevalt suurenenud notsitseptori tundlikkus. 19 20 NGF ja ka teised põletikulised vahendajad sensibiliseerivad TRPV1 sekundaarsete messenger-radade mitmekesise hulga kaudu. Arvatakse, et perifeersete notsitseptorite tundlikkuses osalevad ka paljud teised retseptorid, sealhulgas kolinergilised retseptorid, a-aminovõihappe (GABA) retseptorid ja somatostatiini retseptorid.
Suur hulk põletikulisi vahendajaid on konkreetselt seotud õlavalu ja pöörleva mansetihaigusega. 21 Kuigi mõned keemilised vahendajad aktiveerivad otseselt notsitseptoreid, viivad enamus sensoorses neuronis endasse, selle asemel, et seda otseselt aktiveerida. Need muutused võivad olla varajased translatsioonijärgsed või hilinenud transkriptsioonid. Esimeste näited on membraaniga seotud valkude fosforüülimisest tulenevad muutused TRPV25 retseptoris või pingega seotud ioonkanalites. Viimase näited hõlmavad NGF-indutseeritud TRV1-kanalite tootmise kasvu ja rakusiseste transkriptsioonifaktorite kaltsiumist põhjustatud aktivatsiooni.
Molekulaarsed mehhanismid nakkuse levikust
Valu tunne annab meile märku reaalsest või eelseisvast vigastusest ja käivitab asjakohased kaitsereaktsioonid. Kahjuks ületab valu sageli oma kasulikkuse hoiatussüsteemina ning muutub krooniliseks ja nõrgemaks. See üleminek kroonilisele faasile hõlmab muutusi seljaajus ja ajus, kuid on ka märkimisväärne modulatsioon, kus valusõnumid algatatakse primaarse sensoorse neuroni tasandil. Püüded teha kindlaks, kuidas need neuronid tuvastavad termilist, mehaanilist või keemilist laadi valu tekitavaid stiimuleid, on paljastanud uued signaalimehhanismid ja viinud meid lähemale molekulaarsete sündmuste mõistmisele, mis hõlbustavad üleminekut ägedalt püsivale valule.
Nociceptoride neurokeemia
Glutamaat on domineeriv põletikuline neurotransmitter kõikides nakkõrgendites. Täiskasvanud DRG histokeemilised uuringud näitavad siiski kahte laialdast mittemeliniseeritud C-kiudaineid.
Keemilised muundurid, mis muudavad valu hullemaks
Nagu ülalpool kirjeldatud, suurendab kehavigastus meie valu tekkimist, suurendades nutsütopeptorite tundlikkust nii termiliste kui ka mehaaniliste ärritavate näitajate suhtes. See nähtus tuleneb osaliselt keskkonnamõjude 36 (joonis 3) esmasest sensoorsest terminalist ja mitteneuraalsetest rakkudest (näiteks fibroblastid, nuumrakud, neutrofiilid ja vereliistakud) keemiliste vahendajate tootmisel ja vabanemisel. Mõned põletikupiibi komponendid (näiteks prootonid, ATP, serotoniin või lipiidid) võivad nutsütoprodukti pinnale mõjutada neuronaalset ärritust otseselt, samas kui teised (näiteks bradükiniin ja NGF) seonduvad metabotroopsete retseptoritega ja vahendavad nende mõju läbi sekundaarse signaaliülekande cascades11. Selliste modulaatorsete mehhanismide biokeemiliste aluste mõistmisel on saavutatud märkimisväärset edu.
Rakuvälised prootonid ja kudede atsidoos
Kohalik koeatsidoos on vigastuste füsioloogiline vastus ja kaasnev valu või ebamugavustunne on korreleerunud hapestumise suurusega 37. Happe (pH 5) kasutamine nahal põhjustab püsiva väljalaskmise kolmandas või mitmes polymodal-natsitseptorites, mis indutseerivad vastuvõtlikku välja 20.
Rakulised ja molekulaarsed valumehhanismid
Abstraktne
Närvisüsteem tuvastab ja tõlgendab paljusid soojus- ja mehaanilisi ärritusi, samuti keskkonna- ja endogeenseid keemilisi ärritajaid. Kui need on intensiivsed, põhjustavad need ärritused ägedat valu ja püsiva kehavigastuse korral on valu valgendamise raja nii perifeerse kui ka kesknärvisüsteemi komponendid tohutu plastilisus, valusignaalide paranemine ja ülitundlikkuse tekitamine. Kui plastilisus hõlbustab kaitsvaid reflekse, võib see olla kasulik, kuid kui muutused püsivad, võib tekkida krooniline valu. Geneetilised, elektrofüsioloogilised ja farmakoloogilised uuringud selgitavad molekulaarseid mehhanisme, mis on aluseks valu tekitavate kahjulike stiimulite avastamisele, kodeerimisele ja modulatsioonile.
Sissejuhatus: äge versus püsiv valu
Joonis 5. Seljaaju (keskne) sensibiliseerimine
Glutamaadi / NMDA retseptori vahendatud sensibiliseerimine.Pärast intensiivset stimulatsiooni või püsivat vigastust on aktiveeritud C ja A? notsitseptorid vabastavad mitmesugused neurotransmitterid, sealhulgas dlutamaat, aine P, kaltsitoniinigeeniga seotud peptiid (CGRP) ja ATP, pindmise selja sarve I kihi väljundnärvidele (punane). Selle tagajärjel võivad postsünaptilises neuronis asuvad tavaliselt vaiksed NMDA glutamaadi retseptorid signaali anda, suurendada rakusisest kaltsiumi ja aktiveerida paljusid kaltsiumist sõltuvaid signaaliradasid ja teisi käskjalasid, sealhulgas mitogeeniga aktiveeritud valgukinaasi (MAPK), valgukinaas C (PKC) , proteiinkinaas A (PKA) ja Src. See sündmuste kaskaad suurendab väljundnärvi erutatavust ja hõlbustab valusõnumite edastamist ajju.
Disinhibition.Normaalsetes tingimustes vabastavad inhibeerivad interneuroonid (sinised) pidevalt GABA-d ja / või glütsiini (Gly), et vähendada lamina I väljundnärvide erutatavust ja moduleerida valu ülekannet (inhibeeriv toon). Kuid vigastuse korral võib see pärssimine kaotsi minna, mille tulemuseks on hüperalgeesia. Lisaks võib desinhibitsioon võimaldada mitte-notsitseptiivset müeliniseeritud A? esmaseid aferente valude ülekandelülituse ühendamiseks nii, et tavaliselt kahjutuid stiimuleid tajutakse nüüd valulikena. See toimub osaliselt ergastava PKC? interneuronite väljendamine sisekihis II.
Microglial aktiveerimine.Perifeersed närvikahjustused soodustavad ATP ja kemokiinfraktalkiini vabanemist, mis stimuleerivad mikrogliaalrakke. Eelkõige põhjustab purinergiliste, CX3CR1 ja Tolli sarnaste retseptorite aktiveerimine mikroglial (lilla) ajupõhise neurotroofse faktori (BDNF) vabanemist, mis I-kihi väljund neuronitega ekspresseeritud TrkB retseptorite aktiveerimise kaudu soodustab erutuvust ja suurenenud valu vastuseks nii kahjulikule kui ka kahjutule stimulatsioonile (see tähendab hüperalgeesia ja allodüünia). Aktiveeritud mikroglia vabastab ka hulga tsütokiine, näiteks kasvaja nekroosifaktorit? (TNFa), interleukiin-1? ja 6 (IL-1a, IL-6) ning muud tegurid, mis aitavad kaasa tsentraalsele sensibiliseerimisele.
Põletiku keemiline palavik
Perifeerne sensibiliseerimine tuleneb sagedamini põletikuga seotud muutustest närvikiudude keemilises keskkonnas (McMahon et al., 2008). Seega kaasneb koekahjustusega sageli endogeensete tegurite kogunemine, mis vabanevad vigastatud piirkonnas asuvatest või sinna imbunud aktiveeritud notsitseptoritest või mitte-närvirakkudest (sealhulgas nuumrakud, basofiilid, trombotsüüdid, makrofaagid, neutrofiilid, endoteelirakud, keratinotsüüdid ja fibroblastid). Kollektiivselt. need tegurid, mida nimetatakse põletikuliseks supiks, esindavad laia valikut signaalmolekule, sealhulgas neurotransmitterid, peptiidid (aine P, CGRP, bradükiniin), eikosinoidid ja nendega seotud lipiidid (prostaglandiinid, tromboksaanid, leukotrieenid, endokannabinoidid), neurotrofiinid, tsütokiinid ja kemokiinid, samuti rakuvälised proteaasid ja prootonid. Tähelepanuväärselt ekspresseerivad notsitseptorid ühte või mitut rakupinna retseptorit, mis on võimelised ära tundma ja reageerima kõigile nendele põletikueelsetele või proalgeesilistele ainetele (joonis 4). Sellised koostoimed suurendavad närvikiudude erutatavust, suurendades seeläbi selle tundlikkust temperatuuri või puudutuste suhtes.
Kahtlemata on põletikulise valu vähendamiseks kõige tavalisem lähenemisviis põletikupiima komponentide sünteesi või kogunemise inhibeerimine. Selle kõige paremini näitavad mittesteroidsed põletikuvastased ravimid, nagu aspiriin või ibuprofeen, mis vähendavad põletikulist valu ja hüperalgeesia, inhibeerides prostaglandiini sünteesis osalevaid tsüklooksügenaasi (Cox-1 ja Cox-2). Teine lähenemisviis on blokeerida põletikuliste ainete toime notsitseptoril. Siin tuuakse esile näiteid, mis annavad uut ülevaate perifeerse sensibiliseerimise rakulistest mehhanismidest või mis moodustavad põletikulise valu ravimise uute ravistrateegiate aluse.
NGF on ehk kõige paremini tuntud oma rollina neurotroofse tegurina, mis on nõutav emotsionaalse neuroni elulemuse ja arengu tagamiseks embrüogeneesi ajal, kuid täiskasvanu puhul toodetakse NGF ka koekahjustuse tekkeks ja moodustab põletikupiima olulise komponendi (Ritner et al., 2009). Paljude rakuliste sihtmärkide seas toimib NGF otseselt peptiderigiliste C-kiudotsüüptoridena, mis ekspresseerivad kõrge afiinsusega NGF retseptori türosiini kinaasi, TrkA-d, samuti madala afiinsusega neurotrofiini retseptorit, p75 (Chao, 2003, Snider ja McMahon, 1998). NGF toodab sügavat ülitundlikkust soojuse ja mehhaaniliste stiimulite vastu kahe ajaliselt eraldiseisva mehhanismi kaudu. Alguses aktiveerib NGF-TrkA interaktsioon allavoolu signaaliradade, kaasa arvatud fosfolipaas C (PLC), mitogeen-aktiveeritud proteiinkinaas (MAPK) ja fosfoinositiid 3-kinaas (PI3K). Selle tulemuseks on märklaudvalkude funktsionaalne võimendamine perifeerses natsitseptori terminaalis, eriti TRPV1, mis põhjustab rakulise ja käitumusliku kuumustundlikkuse kiiret muutumist (Chuang et al., 2001).
Sõltumata nende pro-notsitseptiivsetest mehhanismidest on neurotrofiini või tsütokiini signaalide sekkumisest saanud peamiseks strateegiaks põletikuliste haiguste või sellest tuleneva valu ohjamine. Peamine lähenemisviis hõlmab NGF või TNF-? toime neutraliseeriva antikehaga. TNF-a puhul on see olnud märkimisväärselt efektiivne paljude autoimmuunhaiguste, sealhulgas reumatoidartriidi ravis, mis on põhjustanud nii koe hävitamise kui ka sellega kaasneva hüperalgeesia dramaatilise vähenemise (Atzeni et al., 2005). Kuna NGF-i peamised toimed täiskasvanud notsitseptoril toimuvad põletiku taustal, on selle lähenemisviisi eeliseks see, et hüperalgeesia väheneb mõjutamata normaalne valu tajumine. Tõepoolest, anti-NGF antikehad on praegu kliinilistes uuringutes põletikuliste valu sündroomide raviks (Hefti et al., 2006).
Glutamaadi / NMDA retseptori poolt vahendatud sensibiliseerimine
Akuutne valu märgitakse glutamaadi vabastamisest natsitseptorite keskmistest klemmidest, tekitades erutusjärgseid post-sünaptilisi vooge (EPSC-d) teise järjekorra seljaaju neuronites. See toimub peamiselt postsünaptiliste AMPA aktiveerimise ja ioonotroopsete glutamaadi retseptorite alamtüüpide kaudu. Allservpunkti EPSC-de summeerimine postsünaptilise neuronis toob endaga lõpuks kaasa valu-sõnumi edastamise ja kõrgema järjestusega neuronite edasikandumise.
Teised uuringud näitavad, et projektsiooni neuroni muutused ise soodustavad inhibeerivat protsessi. Näiteks reguleerib perifeersete närvikahjustus K + - Cl-kaasaskandur KCC2, mis on oluline normaalsete K + ja Cl-gradientide säilitamiseks kogu plasmamembraanile (Coull et al., 2003). Lamina I projektsiooniga neuronites ekspresseeritud KCC2i alandav regressioon tagab Cl-gradiendi nihke, nii et GABA-A retseptorite aktiveerimine depolariseerub pigem pigem kui lamina I projektsiooni neuronite hüperpolariseerumine. See omakorda suurendaks põletikuvõimet ja suurendaks valuülekannet. Tõepoolest, KCC2i farmakoloogiline blokaad või siRNA vahendatud roti indutseerimine põhjustab mehaanilist allodüüniat.
Jaga ebook
Allikad:
Miks mu õla haiget teeb? Õnguvalu neuroanatoomilise ja biokeemilise baasi ülevaade
Benjamin John Floyd Dean, Stephen Edward Gwilym, Andrew Jonathan Carr
Raua ja molekulaarsed valu mehhanismid
Allan I. Basbaum1, Diana M. Bautista2, Greyory Scherrer1 ja David Julius3
1 Anatoomia osakond, California Ülikool, San Francisco 94158
2Molekulaarsete ja rakubioloogiateaduskond, California Ülikool, Berkeley CA 94720 3Kalifornia Ülikooli füsioloogia osakond, San Francisco 94158
Notsitseptsiooni molekulaarsed mehhanismid
David Julius * ja Allan I. Basbaum
* Raku- ja molekulaarfarmakoloogia osakond ning anatoomia ja füsioloogia osakond ning WM Kecki sihtasutuse integreeriva neuroteaduse keskus, California Ülikool San Francisco, San Francisco, California 94143, USA (e-post: julius@socrates.ucsf.edu)
Neurogeenne põletik, või NI on füsioloogiline protsess, kus vahendajad otse naha närvidest põletikuvastase reaktsiooni alustamiseks. Selle tagajärjel tekivad kohalikud põletikulised reaktsioonid, sealhulgas erüteem, paistetus, temperatuuri tõus, hellus ja valu. Peamised unmeelinitud aferentsed somaatilised C-kiud, mis reageerivad madala intensiivsusega mehaanilistele ja keemilistele stimulatsioonidele, on suuresti vastutavad nende põletikuliste vahendajate vabastamise eest.
Stimuleerituna vabanevad need närvirakud naha närvides energeetilised neuropeptiidid või aine P ja kaltsitoniini geeni seotud peptiid (CGRP) kiiresti mikrokeskkonda, põhjustades põletikuliste reaktsioonide seeria. Immunogeenses põletikus on märkimisväärne erinevus, mis on kõige esimene kaitse- ja reparatiivne vastus, mille immuunsüsteem tekitab, kui patogeen siseneb kehasse, samas kui neurogeensed põletikud hõlmavad otsest seost närvisüsteemi ja põletikuliste reaktsioonide vahel. Kuigi neurogeenne põletik ja immunoloogiline põletik võivad eksisteerida samaaegselt, ei ole need kaks kliiniliselt eristatav. Allpool oleva artikli eesmärk on arutada neurogeense põletiku ja perifeerse närvisüsteemi rolli peremeesorganismi kaitses ja immunopatoloogias.
Neurogeenne põletik - perifeerse närvisüsteemi roll peremeesorganismi kaitses ja immunopatoloogias
Abstraktne
Perifeerset närvisüsteemi ja immuunsüsteemi peetakse tavaliselt eraldi funktsioonideks. Kuid uus joonistamine neurogeense põletiku kohta muutub sellel joonel üha ähmasemaks. Nutsüptor-neuronitel on palju molekulaarset äratundmissuunda, mis on ohuks immuunrakudena ja reageerides ohule, perifeerne närvisüsteem otseselt immuunsüsteemiga, moodustades integreeritud kaitsemehhanismi. Perifeersete kudede mehaaniliste ja autonoomsetest kiududest koosnev nõrk innervatsioonivõrk ja närviülekande kiire kiirus võimaldab kiiret lokaalse ja süsteemse immuunsuse neurogeense modulatsiooni. Näib, et perifeersed neuronid mängivad olulist rolli ka autoimmuunsete ja allergiliste haiguste korral immuunpuudulikkuses. Seetõttu võib perifeersete neuronite ja immuunrakkude kooskõlastatud vastasmõju mõistmine edendada terapeutilisi lähenemisi, et suurendada peremeesorganismi kaitset ja pärssida immunopatoloogiat.
Sissejuhatus
Kaks tuhat aastat tagasi määratles Celsus põletikku nelja kardinaalse nähuna - Dolor (valu), Calor (kuumus), Rubor (punetus) ja kasvaja (turse) - tähelepanek, mis näitas, et närvisüsteemi aktiveerimine tunnistati lahutamatuks osaks põletik. Kuid sellest ajast peale on valu peamiselt mõeldud ainult sümptomina, mitte põletiku tekkimise osaliseks. Selles perspektiivis näitame, et perifeerne närvisüsteem mängib otsest ja aktiivset rolli kaasasündinud ja adaptiivse immuunsuse moduleerimisel, nii et immuunsusel ja närvisüsteemil võib olla peremehe kaitses ühine integreeritud kaitsefunktsioon ja reaktsioon koekahjustusele, mis on keeruline vastasmõju, mis võib põhjustada ka allergiliste ja autoimmuunhaiguste patoloogiat.
Organismide ellujäämine sõltub kriitiliselt võimest luua kaitse koekahjustuste ja nakkuste võimaliku kahju eest. Peremeesorganismi kaitse hõlmab nii vältimist, et eemaldada kontakt ohtliku (kahjuliku) keskkonnaga (närvifunktsioon), kui ka patogeenide aktiivset neutraliseerimist (immuunfunktsioon). Traditsiooniliselt on immuunsüsteemi rolli nakkusetekitajate vastu võitlemisel ja koekahjustuste parandamisel peetud üsna erinevaks närvisüsteemi omast, mis muudab aistingute ja reflekside tekitamiseks kahjulikud keskkonna- ja sisemised signaalid elektriliseks tegevuseks (joonis 1). Teeme ettepaneku, et need kaks süsteemi on tegelikult ühtse kaitsemehhanismi komponendid. Somatosensoorne närvisüsteem on ideaalses kohas ohu avastamiseks. Esiteks, kõiki väliskeskkonnaga tugevalt kokku puutuvaid kudesid, nagu naha, kopsude, kuseteede ja seedetrakti epiteelipinnad, innotseerivad tihedalt notsitseptorid, kõrge lävega valu tekitavad sensoorsed kiud. Teiseks toimub kahjulike väliste stiimulite ülekandmine peaaegu silmapilkselt, suurusjärkude võrra kiiremini kui kaasasündinud immuunsüsteemi mobiliseerimine ja võib seetõttu olla peremeesorganismi kaitses esimene reageerija.
Joonis 1: Mürgised stiimulid, mikroobsed ja põletikulised äratundmissuundid käivitavad perifeerse närvisüsteemi aktiveerimise. Sensori neuronitel on mürgiste / kahjulike stiimulite tuvastamiseks mitu võimalust. 1). Häire signaali retseptorid, sealhulgas TRP kanalid, P2X kanalid ja ohuga seotud molekulaarse mustri (DAMP) retseptorid tunnevad ära keskkonna eksogeensed signaalid (nt kuumus, happesus, kemikaalid) või endogeensed ohumärgid, mis vabanevad trauma / koekahjustuse ajal (nt ATP, kusihape, hüdroksünovaadid). 2) Mudilaskeemi retseptorid (PRR) nagu Toll-sarnased retseptorid (TLR-d) ja Nod-sarnased retseptorid (NLR-id) tunnustavad infektsioonide käigus sissetungivate bakterite või viiruste kaudu nakatatud bakterite või viirustega nakatatud patogeenidega seotud molekulaarseid mustreid (PAMPs). 3) Tsütokiini retseptorid tunnevad ära immuunrakkude (nt IL-1beta, TNF-alfa, NGF) poolt sekreteeritavad tegurid, mis aktiveerivad kaardi kinaasid ja muud signaalimismehhanismid membraani erutatavuse suurendamiseks.
Lisaks perifeeriast pärinevatele seljaaju ja aju ortodroomsetele sisenditele võib notsitseptori neuronite toimepotentsiaalid levida antidrommiliselt ka hargnemispunktides tagasi perifeeriasse, aksoni refleks. Need koos püsivate lokaalsete depolarisatsioonidega põhjustavad närvivahendajate kiiret ja lokaalset vabanemist nii perifeersetest aksonitest kui ka terminalidest (joonis 2) 1. Goltzi (aastal 1874) ja Baylissi (aastal 1901) klassikalised katsed näitasid, et elektriliselt stimuleerivad seljajuured indutseerib naha vasodilatatsiooni, mis viis immuunsüsteemi tekitatud neurogeense põletiku kontseptsioonini (joonis 3).
Joonis 2: Notsüptor-sensoorsed neuronid vabanevad neuronaalsed tegurid põhjustavad otseselt leukotsüütide kemotaksist, vaskulaarset hemodünaamikat ja immuunvastust. Kui mürgised ärritused aktiveerivad sensoorsetes närvides afereeritavaid signaale, genereeritakse antidromic akson refleksid, mis indutseerivad neuropeptiidide vabanemist neuronite perifeersetes terminalides. Neil molekulaarsetel mediaatoritel on mitmeid põletikulisi toimemehhanisme: 1). Kemotaksis ja neutrofiilide, makrofaagide ja lümfotsüütide aktivatsioon kahjustuskohta ja nuumrakkude degranulatsioon. 2) Vaskulaarsete endoteliaalsete rakkude signalisatsioon verevoolu, vaskulaarse lekke ja ödeemi tekkeks. See võimaldab ka põletikuliste leukotsüütide värbamist. 3) Dendriitrakkude ristamine järgneva T-abistajarakkude diferentseerumise juhtimiseks Th2-i või Th17-i alamtüüpideks.
Joonis 3: Celsuse põletiku neurogeensete aspektide mõistmise arenemise ajajärk tänapäevani.
Neurogeenset põletikku vahendab neuropeptiidide kaltsitoniini geeniga seotud peptiidi (CGRP) ja substantsi P (SP) vabanemine notsitseptoritest, mis toimivad otseselt veresoonte endoteeli- ja silelihasrakkudele 2. CGRP tekitab vasodilatatsiooniefekte 5, 2, samas kui SP suurendab kapillaaride läbilaskvust, mis põhjustab plasma ekstravasatsiooni ja turset 3, 4, aidates kaasa Celsuse hõõrdumise, kalorite ja kasvaja tekkele. Kuid notsitseptorid vabastavad palju täiendavaid neuropeptiide (veebibaas: www.neuropeptides.nl/), sealhulgas adrenomedulliin, neurokiniinid A ja B, vasoaktiivne soolepeptiid (VIP), neuropeptiid (NPY) ja gastriini vabastav peptiid (GRP), samuti muud molekulaarsed vahendajad nagu glutamaat, lämmastikoksiid (NO) ja tsütokiinid nagu eotaksiin 6.
Nüüd mõistame, et perifeeria sensoorsetest neuronitest vabanenud vahendajad mitte ainult ei mõjuta veresooni, vaid meelitavad ja aktiveerivad otseselt ka kaasasündinud immuunrakke (nuumrakud, dendriitrakud) ja adaptiivseid immuunrakke (T-lümfotsüüdid) 7. Koekahjustuse ägedas olukorras oletame, et neurogeenne põletik on kaitsev, hõlbustades immuunrakkude aktiveerimise ja värbamise abil haavade füsioloogilist paranemist ja immuunsust patogeenide eest. Kuid sellistel neuro-immuunsustel on tõenäoliselt ka suur roll allergiliste ja autoimmuunhaiguste patofüsioloogias, võimendades patoloogilisi või kohanemisvastaseid immuunvastuseid. Näiteks reumatoidartriidi loommudelites on Levine ja tema kolleegid näidanud, et liigese denervatsioon põhjustab silmatorkavat põletiku nõrgenemist, mis sõltub aine P 12, 13 närvilisest ekspressioonist. Hiljutistes uuringutes allergilise hingamisteede põletiku, koliidi ja psoriaasi korral on primaarsetel sensoorsetel neuronitel keskne roll kaasasündinud ja adaptiivse immuunsuse aktiveerimisel ja suurendamisel.
Seega pakume välja, et perifeerne närvisüsteem ei osale mitte ainult passiivses rollis peremeesorganismi kaitsmises (kahjulike stiimulite avastamine ja vältiva käitumise algatamine), vaid ka aktiivne roll koos immuunsüsteemiga kahjulike stiimulid, roll, mida saab haiguste tekkeks panna.
Perifeersete närvide ja eluvõimeliste immuunsüsteemide ühised ohutegurid
Perifeersed sensoorsed neuronid on kohandatud tunnetama organismi ohtu nende tundlikkuse tõttu intensiivsetele mehhaanilistele, termilistele ja ärritavatele keemilistele stiimulitele (joonis 1). Transientne retseptori potentsiaal (TRP) ioonkanalid on kõige levinumad mittespetsiifilised molekulaarsed mediaatorid, mis juhivad mittenektiivset katioonide sisenemist mitmete mürgiste stiimulite aktiveerimisel. TRPV1 aktiveeritakse kõrgete temperatuuride, madala pH-ga ja kapsaitsiiniga, chilipipartide 18 vallinoidärritava komponendiga. TRPA1 vahendab reaktiivsete kemikaalide, sealhulgas keskkonna ärritavate ainete nagu pisargaasi ja tööstuslike isotiotsüanaatide 19 tuvastamist, kuid mis veelgi olulisem, see aktiveerub ka koekahjustuse ajal endogeensete molekulaarsete signaalide kaudu, sealhulgas 4-hüdroksünonenaal ja prostaglandiinid 20, 21.
Huvitav on see, et sensoorsed neuronid omavad paljudel samadel patogeenidel ja ohu molekulaarse äratundmise retseptoriteede kui sisemistest immuunrakudest, mis võimaldavad neil ka tuvastada patogeene (joonis 1). Immuunsüsteemis tuvastatakse mikroobsete patogeenide abil iduliini poolt kodeeritud retseptorid (PRR), mis tunnustavad üldjoontes konserveerunud eksogeenseid patogeenidega seotud molekulaarseid mustreid (PAMP-sid). Esimesed PRR-id, mis olid identifitseeritud, olid teemaksarnase retseptori (TLR) perekonna liikmed, mis seostuvad pärmi, bakteriaalsetest rakuseina komponentidest ja viiruse RNA 22-st. Pärast PRR-i aktiveerimist lülitatakse sisse allavoolu signaalirajad, mis indutseerivad tsütokiini tootmist ja aktiveerib adaptiivset immuunsust. Lisaks TLR-ile aktiveeritakse ka sisemine immune rakke koekahjustuse ajal endogeensed tuletatud ohusignaalid, mida nimetatakse ka kahjustatud molekulaarseteks mustriteks (DAMPs) või alarmins 23, 24. Need ohusignaalid hõlmavad HMGB1, kusihapet ja kuumašoki valgud, mis vabanevad surevate rakkude ajal nekroosi ajal, aktiveerides immuunrakud nakatamatute põletikuvastaste reaktsioonide ajal.
PRR-sid, sealhulgas TLR-sid 3, 4, 7 ja 9, ekspresseerivad notsitseptorite neuronid ning stimulatsioon TLR-ligandidega viib sissepoole suunatud voolude indutseerimiseni ja notsitseptorite sensibiliseerimiseni teiste valu stiimulite suhtes 25, 27. Lisaks viib sensoorsete neuronite aktiveerimine TLR7 ligandi imikvimoodi poolt sügelusspetsiifilise sensoorsuse raja 25 aktiveerimiseni. Need tulemused näitavad, et infektsiooniga seotud valu ja sügelus võivad osaliselt olla tingitud neuronite otsesest aktiveerimisest patogeenist tulenevate tegurite poolt, mis omakorda aktiveerida immuunrakke neuronaalsete signaalmolekulide perifeerse vabastamise kaudu.
Peamine rakukahjustuse ajal vabanev DAMP / alariin on ATP, mille tunnevad ära nii notsitseptori neuronite kui ka immuunrakkude purinergilised retseptorid 28-30. Purinergilised retseptorid koosnevad kahest perekonnast: P2X retseptorid, ligandiga seotud katioonikanalid ja P2Y retseptorid, G-valguga seotud retseptorid. Notsitseptor-neuronites toimub ATP äratundmine P2X3 kaudu, mis viib kiiresti tihedustundvate katioonivoolude ja valu 28, 30 (joonis 1), samas kui P2Y retseptorid aitavad kaasa notsitseptori aktiveerimisele TRP ja pingega seotud naatriumikanalite sensibiliseerimise kaudu. Makrofaagides põhjustab ATP seondumine P2X7 retseptoritega hüperpolarisatsiooni ja IL-1 beeta ja IL-18 29 genereerimisel olulise molekulaarse kompleksi, põletikulise molekuli aktiveerimise. Seetõttu on ATP tugev ohusignaal, mis aktiveerib nii perifeersed neuronid kui ka kaasasündinud immuunsus vigastuse ajal ja mõned tõendid viitavad isegi sellele, et neuronid ekspresseerivad põletikulise molekulaarse masina osi 31.
Notsitseptorites esinevate ohtlike signaalide pöörde külg on TRP-kanalite roll immuunrakkude aktiveerimisel. TRPV2, mürgise kuumusega aktiveeritud TRPV1 homoloog, ekspresseeritakse sünnieelsete immuunrakkude 32 kõrgel tasemel. TRPV2i geneetiline ablatsioon viis makrofaagide fagotsütoosi ja bakteriaalsete infektsioonide 32 kliirensini. Nuumrakud ekspresseerivad ka TRPV kanaleid, mis võivad otseselt vahendada nende degranulatsiooni 33. Jääb kindlaks teha, kas endogeensed ohusignaalid aktiveerivad immuunkrele sarnaselt notsitseptoritega.
Immuunrakkude ja notsitseptor-neuronite peamine sidevahend on tsütokiinide kaudu. Tsütokiiniretseptorite aktiveerimisel aktiveeritakse sensoorsetes neuronites signaaliülekande teed, mis viib membraanivalkude, sealhulgas TRP ja pingega kanalite fosforüülimiseni allavoolu (joonis 1). Sellest tulenev notsitseptorite sensibiliseerimine tähendab, et tavaliselt võivad kahjutud mehaanilised ja soojuse stiimulid nüüd aktiveerida notsitsepte. Interleukiin 1 beeta ja TNF-alfa on kaks olulist tsütokiini, mille põletikulised immuunrakud vabastavad. IL-1 beeta ja TNF-alfa tajuvad otseselt notsitseptorid, mis ekspresseerivad sugulasretseptoreid, indutseerivad p38 kaardikinaaside aktivatsiooni, mis viib membraani suurenenud erutuvuse 34. Närvikasvufaktor (NGF) ja prostaglandiin E (36) on samuti peamised immuunrakkudest vabanevad põletikulised vahendajad, mis toimivad sensibiliseerivalt otseselt perifeersetel sensoorsetel neuronitel. Notsitseptorite sensibiliseerimise oluline mõju immuunfaktorite poolt on neuropeptiidide suurenenud vabanemine perifeersetes terminalides, mis veelgi aktiveerivad immuunrakke, kutsudes seeläbi esile positiivse tagasiside, mis juhib ja hõlbustab põletikku.
Sensorne närvisüsteemi kontrollimine intuitiivse ja adaptiivse immuunsuse vastu
Põletiku varajastes faasides annavad sensoorsed neuronid signaali koes elavatele nuumrakkudele ja dendriitrakkudele, mis on immuunvastuse algatamisel olulised kaasasündinud immuunrakud (joonis 2). Anatoomilised uuringud on näidanud terminalide otsest paiknemist nuumrakkudega, samuti dendriitrakkudega ning notsitseptoritest vabanevad neuropeptiidid võivad nendes rakkudes 7, 9, 37 esile kutsuda degranulatsiooni või tsütokiinide tootmist. Sellel koostoimel on allergiliste hingamisteede puhul oluline roll põletik ja dermatiit 10.
Põletiku efektorfaasis peavad immuunrakud leidma tee konkreetsesse vigastuskohta. Paljud sensoorsetest neuronitest, neuropeptiididest, kemokiinidest ja glutamaadist vabanenud vahendajad on neutrofiilide, eosinofiilide, makrofaagide ja T-rakkude jaoks kemotaktilised ning parandavad endoteeli adhesiooni, mis hõlbustab immuunrakkude kodustamist 6, 38, 41 (joonis 2). Lisaks viitavad mõned tõendid sellele, et neuronid võivad otseselt osaleda efektorfaasis, kuna neuropeptiididel endil võivad olla otsesed antimikroobsed funktsioonid 42.
Neuronaalselt saadud signaalmolekulid võivad suunata ka põletiku tüüpi, aidates kaasa erinevat tüüpi adaptiivsete immuun-T-rakkude diferentseerumisele või spetsifikatsioonile. Antigeeni fagotsütoosivad ja töötlevad kaasasündinud immuunrakud, mis seejärel migreeruvad lähimasse lümfisõlme ja esitavad antigeense peptiidi naistele T-rakkudele. Sõltuvalt antigeeni tüübist, kaasasündinud immuunrakkude kostimuleerivatest molekulidest ja spetsiifiliste tsütokiinide kombinatsioonidest küpsevad vanad T-rakud spetsiifilisteks alatüüpideks, mis kõige paremini teenivad patogeense stiimuli puhastamiseks vajalikke põletikulisi jõupingutusi. CD4 T-rakud või T-abistaja (Th) rakud võib jagada nelja põhirühma: Th1, Th2, Th17 ja T regulatoorsed rakud (Treg). Th1 rakud on seotud peamiselt rakusiseste mikroorganismide ja elundispetsiifiliste autoimmuunhaiguste immuunvastuse reguleerimisega; Th2 on rakuväliste patogeenide, näiteks helmintide suhtes immuunsuse seisukohalt kriitilise tähtsusega ja vastutab allergiliste põletikuliste haiguste eest; Th17 rakkudel on keskne roll kaitsmisel mikroobide, näiteks rakuväliste bakterite ja seente eest; Tregi rakud on seotud enesetaluvuse säilitamise ja immuunvastuste reguleerimisega. Tundub, et seda T-rakkude küpsemisprotsessi mõjutavad sensoorsed neuronaalsed vahendajad. Neuropeptiidid, nagu CGRP ja VIP, võivad dendriitrakke kallutada Th2-tüüpi immuunsuse poole ja vähendada Th1-tüüpi immuunsust, soodustades teatud tsütokiinide tootmist ja pidurdades teisi, samuti vähendades või tõhustades dendriitrakkude migratsiooni kohalikesse lümfisõlmedesse 8 , 10, 43. Sensoorsed neuronid aitavad märkimisväärselt kaasa ka allergilisele (peamiselt Th2-ajendatud) põletikule 17. Lisaks Th1- ja Th2-rakkude reguleerimisele võivad ka teised neuropeptiidid, näiteks SP ja Hemokinin-1, põletikulist vastust rohkem juhtida Th17 või Tregi suunas 44, 45, mis tähendab, et neuronid võivad olla seotud ka põletikulise eraldatuse reguleerimisega. Selliste immunopatoloogiate korral nagu koliit ja psoriaas võib neuronaalsete vahendajate nagu aine P blokeerimine märkimisväärselt summutada T-rakke ja immuunvahendatud kahjustusi 15, ehkki ühe vahendaja antagoniseerimisel võib iseenesest olla neurogeensele põletikule ainult piiratud mõju.
Arvestades, et perifeersetest sensoorsetest närvikiududest vabanenud signaalimolekulid reguleerivad mitte ainult väikesi veresooni, vaid ka kemotaksist, isandamist, küpsemist ja aktiveerimist immuunrakkudes, on selge, et neuroimmuunsed interaktsioonid on palju keerukamad kui varem arvati (joonis 2). Pealegi on üsna mõeldav, et see ei ole individuaalsed neuronid vahendajad, vaid pigem spetsiifilised nitsitseptorid vabanevad signaalimolekulide kombinatsioonid, mis mõjutavad immuunvastuse erinevaid astmeid ja tüüpe.
Immuunsuse autonoomne kontrollfunktsioon
Kolinergilise autonoomse närvisüsteemi "refleksi" ahela roll perifeerse immuunvastuse reguleerimisel näib samuti silmapaistev 46. Vagus on peamine parasümpaatiline närv, mis ühendab ajutüve siseorganitega. Kevin Tracey ja teiste töö osutab septilise šoki ja endotokseemia tugevatele generaliseerunud põletikuvastastele reaktsioonidele, mille vallandab ebanormaalne vagaalse närvi aktiivsus, mis viib perifeersete makrofaagide supressioonini 47. Vagus aktiveerib põrna innerveerivaid perifeerseid adrenergilisi tsöliaakiaganglioni neuroneid, mis viib atsetüülkoliini allavoolu eraldumiseni, mis seondub põrna ja seedetrakti makrofaagide alfa-49 nikotiiniretseptoritega. See kutsub esile JAK7 / STAT2 SOCS3 signaaliraja aktiveerimise, mis pärsib võimsalt TNF-alfa transkriptsiooni 3. Adrenergiline tsöliaakiaganglion suhtleb otseselt ka atsetüülkoliini tootvate mälu T-rakkude alamhulgaga, mis pärsivad põletikulisi makrofaage 47.
Invaratsionaalsed looduslikud Killer-T-rakud (iNKT) on spetsiifiline T-rakkude alamhulk, mis peptiidi antigeenide asemel tuvastab mikroobide lipiidid CD1d kontekstis. NKT rakud on peamine lümfotsüütide populatsioon, kes osaleb nakkushaiguste vastases võitluses ja süsteemse immuunsuse reguleerimises. NKT rakud elavad ja liikuvad põhiliselt põrna ja maksa veresoonte ja sinusioidide kaudu. Maksa sümpaatilised beeta-adrenergilised närvid kinnitavad otseselt NKT rakkude aktiivsuse 50 modulatsiooni. Hoolimata hiire mudelist (MCAO), näiteks NKT-rakkude mobiilsus oli nähtavalt allasurutud, mis oli pööratud sümpaatilise denervatsiooni või beeta-adrenergiliste antagonistide poolt. Veelgi enam, NAD-rakkude noradrenergiliste neuronite immunosupressiivne aktiivsus tõi kaasa süsteemse nakkuse ja kopsukahjustuse suurenemise. Seetõttu võivad autonoomsete neuronite efektorne signaalid vahendada tugevat immuunvastust.
Dr Alex Jimenez Insight
Neurogeenne põletik on närvisüsteemi tekitatud lokaalne põletikuvastane reaktsioon. Arvatakse, et see mängib olulist rolli mitmesuguste terviseprobleemide, sealhulgas migreeni, psoriaasi, astma, fibromüalgia, ekseemi, rosaatsea, düstoonia ja mitmete keemiliste tundlikkuste patogeneesis. Kuigi kaugelearenenud närvisüsteemiga seotud neurogeenset põletikku on põhjalikult uuritud, vajab kesknärvisüsteemi neurogeense põletiku mõiste edasist uurimist. Mitmete uurimistööde kohaselt on neurogeense põletiku peamine põhjus magneesiumipuudus. Järgnev artikkel annab ülevaate närvisüsteemi neurogeense põletiku mehhanismidest, mis võivad aidata tervishoiutöötajatel määrata parima ravivõimaluse, et hoolitseda erinevate närvisüsteemiga seotud terviseprobleemide eest.
Järeldused
Millised on somatosensoorse ja autonoomse närvisüsteemi spetsiifilised rollid põletiku ja immuunsüsteemi reguleerimisel (joonis 4)? Notsitseptorite aktiveerimine viib lokaalsete aksonirefleksideni, mis värbavad ja aktiveerivad lokaalselt immuunrakke ning on seetõttu peamiselt põletikku soodustavad ja ruumiliselt piiratud. Seevastu autonoomne stimulatsioon viib süsteemse immunosupressioonini, mõjutades maksa ja põrna immuunrakkude kogumeid. Perifeeria aferentsed signaalimehhanismid, mis põhjustavad immunosupressiivse vagaalse kolinergilise refleksi ahela käivitamise, on halvasti mõistetavad. 80–90% vagulaarsetest kiududest on siiski primaarsed aferentsed sensoorsed kiud ja seetõttu võivad siseelunditest pärinevad signaalid, mida paljud võivad mõjutada immuunrakud, viia interneuroonide aktiveerimiseni ajutüves ja nende kaudu väljumiseni efferentsetes vagulaarsetes kiududes 46.
Joonis 4: Sensoorne ja autonoomsed närvisüsteemid moduleerivad vastavalt kohalikke ja süsteemseid immuunvastuseid. Epiteeli pindade (nt nahk ja kopsud) innerveerivad nukleotiidid indutseerivad lokaalseid põletikuvastuseid, nuumrakkude ja dendriitrakkude aktiveerimist. Allergilisel hingamisteede põletikul, dermatiitil ja reumatoidartriidil võivad nutsüptor-neuronid põletikku juhtida. Seevastu reguleerivad süsteemsed immuunvastused makrofaagide ja NKT rakkude aktiveerimise blokeerimisega automaatsed sisselaskeorganite (nt põrna ja maksa) sissetungivad autonoomsed ahelad. Kui insult ja septiline endotokseemia avaldavad neuronid immunosupressiivset rolli.
Tüüpiliselt määratletakse põletiku ajakäik ja olemus, kas infektsiooni ajal, allergiliste reaktsioonide või autoimmuunpatoloogia korral, kaasatud immuunrakkude kategooriate järgi. Oluline on teada, milliseid erinevaid immuunrakke reguleerivad sensoorsed ja autonoomsed signaalid. See küsimus võiks lahendada süstemaatilise hinnangu andmisest selle kohta, milliseid vahendajaid saab vabastada notsitseptoritest ja autonoomsetest neuronitest ning nende poolt retseptorite ekspressioon erinevatest sünnilistest ja adaptiivsetest immuunrakkudest.
Evolutsiooni käigus on nii kaasasündinud immuunsuse kui ka notsitseptsiooni jaoks välja töötatud sarnased ohu tuvastamise molekulaarsed teed, kuigi rakkudel on täiesti erinevad arenguliinid. Kui immunoloogid ja neurobioloogid uurivad PRR-sid ja kahjulike ligandiga seotud ioonkanaleid eraldi, siis piir nende kahe välja vahel on üha hägusam. Koekahjustuste ja patogeense infektsiooni ajal viib ohusignaalide vabanemine tõenäoliselt nii perifeersete neuronite kui ka immuunrakkude kooskõlastatud aktiveerumiseni keeruka kahesuunalise suhtluse ja integreeritud peremeeskaitsega. Notsitseptorite anatoomiline positsioneerumine keskkonnaga kokkupuutel, närviülekande kiirus ja nende võime vabastada immuuntoimega vahendajate tugevaid kokteile võimaldab perifeersel närvisüsteemil aktiivselt moduleerida kaasasündinud immuunvastust ja koordineerida allavoolu adaptiivset immuunsust. Ja vastupidi, notsitseptorid on väga tundlikud immuunvahendajate suhtes, mis aktiveerivad ja sensibiliseerivad neuroneid. Neurogeenne ja immuunvahendatud põletik ei ole seetõttu iseseisvad üksused, vaid toimivad koos varajase hoiatamise seadmetena. Kuid perifeersel närvisüsteemil on oluline roll ka paljude immuunhaiguste, nagu astma, psoriaas või koliit, patofüsioloogias ja võib-olla ka etioloogias, kuna selle võime immuunsüsteemi aktiveerida võib võimendada patoloogilist põletikku 15-17. Immuunsushäirete ravi võib seetõttu hõlmata nii notsitseptorite kui ka immuunrakkude sihtimist.
Tänusõnad
Täname NIH toetuse eest (2R37NS039518).
KokkuvõtteksNeurogeense põletiku rolli mõistmine peremeesorganismi kaitse ja immunopatoloogia osas on väga oluline, et määrata kindlaks õige ravimeetod mitmesuguste närvisüsteemi terviseprobleemide korral. Vaadates perifeersete neuronite koostoimeid immuunrakkudega, võivad tervishoiutöötajad edendada terapeutilisi lähenemisviise, et veelgi suurendada peremehe kaitset ja pärssida immunopatoloogiat. Eespool nimetatud artikli eesmärk on aidata patsientidel mõista neuropaatia kliinilist neurofüsioloogiat muude närvikahjustuste terviseprobleemide hulgas. Riikliku biotehnoloogiaalase teabe keskuse (NCBI) viidatud teave. Meie teabe ulatus piirdub kiropraktika, samuti selgroovigastuste ja seisunditega. Teema arutamiseks küsige palun dr Jimenezilt või võtke meiega ühendust aadressil915-850-0900.
Juhib dr Alex Jimenez
Täiendavad teemad: Seljavalu
Seljavalu on üks kõige levinumaid puudeid ja töölt puuduvaid tööpäevi kogu maailmas. Tegelikkuses on seljavalu olnud ka arstikeskuse külastuste teine kõige levinum põhjus, millest arvukus oli ainult ülemiste hingamisteede nakkustega. Ligikaudu 80 protsenti elanikkonnast kogeb teatud tüüpi seljavalu vähemalt korra kogu eluea jooksul. Lülisamba on keeruline struktuur, mis koosneb teistest pehmetest kudedest koosnevate luude, liigeste, sidemete ja lihaste vahel. Selle tagajärjel tekivad vigastused ja / või raskendavad seisundid, näiteks hernide kettad, võib lõpuks põhjustada seljavalu sümptomeid. Sageli on seljavalu kõige sagedasem spordivigastus või autoõnnetusjuhtumite põhjus, kuid mõnikord võib kõige lihtsam liikumine olla valulike tulemustega. Õnneks võivad alternatiivsed ravivõimalused, nagu kiropraktiline ravi, aidata leevendada seljavalu, kasutades selleks spinaalset reguleerimist ja manuaalseid manipulatsioone, parandades lõpuks valu hõlbustamist.
2. Edvinsson L, Ekman R, Jansen I, McCulloch J, Uddman R. Kaltsitoniini geeniga seotud peptiid ja ajuveresooned: jaotumine ja vasomotoorne toime.J Aju verevool Metab1987;7: 720 728. [PubMed]
3. McCormacki peadirektoraat, Mak JC, Coupe MO, Barnes PJ. Kaltsitoniini geeniga seotud peptiidi vasodilatatsioon inimese kopsuveresoontesJ Appl Physiol1989;67: 1265 1270. [PubMed]
4. Saria A. Sensoorsetes närvikiududes sisalduv aine P aitab pärast termilist vigastust tekkida turse roti tagakäpas.Br J Pharmacol1984;82: 217 222. [PMC tasuta artikkel][PubMed]
5. Aju SD, Williams TJ. Tahhükiniinide ja kaltsitoniiniga genereeritud peptiidi koostoimed põhjustavad turse moodustumise ja verevoolu moduleerimist roti nahas.Br J Pharmacol1989;97: 77-82.[PMC tasuta artikkel][PubMed]
6. Fritter AD jt. Neuronaalne eotaksiin ja CCR3 antagonisti mõju hingamisteede hüperreaktiivsusele ja M2 retseptori düsfunktsioonile.J Clin Invest2006;116: 228 236. [PMC tasuta artikkel][PubMed]
7. Ansel JC, Brown JR, Payani peadirektoraat, Brown MA. Aine P aktiveerib selektiivselt TNF-alfa geeni ekspressiooni hiire nuumrakkudesJ Immunol1993;150: 4478 4485. [PubMed]
9. Hosoi J jt. Langerhansi raku funktsiooni reguleerimine närvidega, mis sisaldavad kaltsitoniini geeniga seotud peptiidiLoodus1993;363: 159 163. [PubMed]
10. Mikami N jt. Kaltsitoniini geeniga seotud peptiid on oluline naha immuunsuse regulaator: mõju dendriitrakkude ja T-rakkude funktsioonidele.J Immunol2011;186: 6886 6893. [PubMed]
11. Rochlitzer S jt. Neuropeptiidkaltsitoniini geeniga seotud peptiid mõjutab dendriitrakkude funktsiooni moduleerides hingamisteede allergilist põletikku.Clin Exp allergia2011;41: 1609 1621. [PubMed]
12. Cyphert JM jt. Koostöö nuumrakkude ja neuronite vahel on antigeeni vahendatud bronhide ahenemise jaoks hädavajalikJ Immunol2009;182: 7430 7439. [PMC tasuta artikkel][PubMed]
13. Levine JD jt. Intraneuronaalne aine P aitab kaasa eksperimentaalse artriidi raskusastmeleTeadus1984;226: 547 549. [PubMed]
14. Levine JD, Khasar SG, Roheline PG. Neurogeenne põletik ja artriitAnn NY Acad Sci2006;1069: 155 167. [PubMed]
15. Engel MA jt. TRPA1 ja aine P vahendavad koliiti hiirtelGastroenteroloogia2011;141: 1346 1358. [PubMed]
16. Ostrowski SM, Belkadi A, Loyd CM, Diaconu D, Ward NL. Psoriasiformse hiire naha naha denervatsioon parandab sensoorsest neuropeptiidist sõltuvalt akantoosi ja põletikku.J Invest Dermatol2011;131: 1530 1538. [PMC tasuta artikkel][PubMed]
17. Caceres AI jt. Sensoorne neuronaalne ioonkanal, mis on oluline hingamisteede põletikul ja astma hüperreaktiivsuselProc Natl Acad Sci USA A.2009;106: 9099 9104. [PMC tasuta artikkel][PubMed]
18. Caterina MJ jt. Notsitseptiivsuse ja valuaistingu halvenemine hiirtel, kellel puudub kapsaitsiini retseptorTeadus2000;288: 306 313. [PubMed]
19. Bessac BF jt. Ajutise retseptori potentsiaalsed anküliini 1 antagonistid blokeerivad toksiliste tööstuslike isotsüanaatide ja pisargaaside kahjuliku mõju.FASEB J. 2009;23: 1102 1114. [PMC tasuta artikkel][PubMed]
20. Cruz-Orengo L jt. 15-delta PGJ2 poolt ioonikanali TRPA1 aktiveerimise kaudu esile kutsutud nahaotsitseptsioonMol Pain2008;4: 30. [PMC tasuta artikkel][PubMed]
21. Trevisani M jt. 4-hüdroksünonenaal, endogeenne aldehüüd, põhjustab ärritava retseptori TRPA1 aktiveerimise kaudu valu ja neurogeenset põletikku.Proc Natl Acad Sci USA A.2007;104: 13519 13524. [PMC tasuta artikkel][PubMed]
22. Janeway CA, Jr, Medzhitov R. Sissejuhatus: kaasasündinud immuunsuse roll adaptiivses immuunvastuses.Semin Immunol1998;10: 349 350. [PubMed]
23. Matzinger P. kaasasündinud ohutunneAnn NY Acad Sci2002;961: 341 342. [PubMed]
24. Bianchi MINA. DAMP-id, PAMP-id ja alarminid: kõik, mida me peame ohu kohta teadmaJ Leukoc Biol2007;81: 1 5. [PubMed]
25. Liu T, Xu ZZ, Park CK, Berta T, Ji RR. Maksulaadne retseptor 7 vahendab sügelustNat Neurosci2010;13: 1460 1462. [PMC tasuta artikkel][PubMed]
26. Diogenes A, Ferraz CC, Akopian AN, Henry MA, Hargreaves KM. LPS sensibiliseerib TRPV1 TLR4 aktiveerimise kaudu kolmiknärvi sensoorsetes neuronitesJ Dent Res2011;90: 759 764. [PubMed]
27. Qi J jt. Seljaaju ganglioni neuronite TLR-stimuleerimisega indutseeritud valulikud rajadJ Immunol2011;186: 6417 6426. [PMC tasuta artikkel][PubMed]
28. Cockayne DA jt. Kusepõie hüporefleksia ja valuga seotud käitumise vähenemine P2X3 puudulikkusega hiirtelLoodus2000;407: 1011 1015. [PubMed]
29. Mariathasan S jt. Krüopüriin aktiveerib põletikulise reaktsiooni toksiinidele ja ATP-leLoodus2006;440: 228 232. [PubMed]
30. Souslova V jt. Soojad kodeerivad defitsiidid ja kõrvalekalduvad põletikulised valud hiirtel, kellel puuduvad P2X3 retseptoridLoodus2000;407: 1015 1017. [PubMed]
31. de Rivero Vaccari JP, Lotocki G, Marcillo AE, Dietrich WD, Keane RW. Neuronite molekulaarne platvorm reguleerib põletikku pärast seljaaju vigastustJ Neurosci2008;28: 3404 3414. [PubMed]
32. Link TM jt. TRPV2-l on makrofaagide osakeste seondumisel ja fagotsütoosis keskne rollNat Immunol2010;11: 232 239. [PMC tasuta artikkel][PubMed]
33. Turner H, del Carmen KA, Stokes A. Seos TRPV kanalite ja nuumrakkude funktsiooni vahelHandb Exp Pharmacol2007: 457 471[PubMed]
34. Binshtok AM jt. Notsitseptorid on interleukiin-1 beetaanduridJ Neurosci2008;28: 14062-14073.[PMC tasuta artikkel][PubMed]
35. Zhang XC, Kainz V, Burstein R, Levy D. Kasvaja nekroosifaktor alfa kutsub esile kohaliku COX ja p38 MAP kinaasi toimel vahendatud meningeaalsete notsitseptorite sensibiliseerimise.Valu2011;152: 140-149.[PMC tasuta artikkel][PubMed]
36. Samad TA jt. Interleukiin-1 beeta-vahendatud Cox-2 induktsioon kesknärvisüsteemis aitab kaasa põletikulise valu ülitundlikkusele.Loodus2001;410: 471 475. [PubMed]
38. Smith CH, Barker JN, Morris RW, MacDonald DM, Lee TH. Neuropeptiidid kutsuvad esile endoteelirakkude adhesioonimolekulide kiire ekspressiooni ja kutsuvad esile inimese naha granulotsüütilise infiltratsiooni.J Immunol1993;151: 3274 3282. [PubMed]
39. Dunzendorfer S, Meierhofer C, Wiedermann CJ. Signalisatsioon inimese eosinofiilide neuropeptiidide põhjustatud migratsioonisJ Leukoc Biol1998;64: 828 834. [PubMed]
40. Ganor Y, Besser M, Ben-Zakay N, Unger T, Levite M. Inimese T-rakud ekspresseerivad funktsionaalset ionotroopset glutamaadi retseptorit GluR3 ja glutamaat käivitab ise integriini vahendatud adhesiooni laminiini ja fibronektiini ning kemotaktilise migratsiooni.J Immunol2003;170: 4362 4372. [PubMed]
41. Czepielewski RS jt. Gastriini vabastav peptiidiretseptor (GRPR) vahendab kemotaksist neutrofiilidesProc Natl Acad Sci USA A.2011;109: 547 552. [PMC tasuta artikkel][PubMed]
42. Brogden KA, Guthmiller JM, Salzet M, Zasloff M. Närvisüsteem ja kaasasündinud immuunsus: neuropeptiidide ühendus.Nat Immunol2005;6: 558 564. [PubMed]
43. Jimeno R jt. VIP mõju tsütokiinide ja aktiveeritud abistajate T-rakkude põhiregulaatorite vahelisele tasakaaluleImmunol Cell Biol2011;90: 178 186. [PubMed]
44. Razavi R jt. TRPV1 + sensoorsed neuronid kontrollivad beeta-rakkude stressi ja saarepõletikku autoimmuunse diabeedi korralLahter2006;127: 1123 1135. [PubMed]
45. Cunin P jt. Tahhükiniinide aine P ja hemokiniin-1 soodustavad inimese mälu Th17 rakkude teket, indutseerides monotsüütide poolt IL-1 beeta, IL-23 ja TNF-laadse 1A ekspressiooni.J Immunol2011;186: 4175 4182. [PubMed]
46. Andersson U, Tracey KJ. Immunoloogilise homöostaasi reflekspõhimõttedAnnu Rev Immunol2011[PMC tasuta artikkel][PubMed]
47. de Jonge WJ jt. Vagusnärvi stimuleerimine nõrgendab makrofaagide aktivatsiooni, aktiveerides Jak2-STAT3 signaaliraja.Nat Immunol2005;6: 844 851. [PubMed]
48. Rosas-Ballina M jt. Atsetüülkoliini sünteesivad T-rakud edastavad närvisignaale vaguse närviringlusesTeadus2011;334: 98 101. [PMC tasuta artikkel][PubMed]
49. Wang H jt. Nikotiinse atsetüülkoliini retseptori alfa7 alaühik on põletiku oluline reguleerijaLoodus2003;421: 384 388. [PubMed]
50. Wong CH, Jenne CN, Lee WY, Leger C, Kubes P. Maksa iNKT rakkude funktsionaalne innervatsioon on insuldi järgselt immunosupressiivne.Teadus2011;334: 101 105. [PubMed]
IFM-i tööriist Leia praktikant on funktsionaalse meditsiini suurim saatevõrgustik, mis on loodud selleks, et aidata patsientidel leida funktsionaalse meditsiini praktikuid kõikjal maailmas. IFMi sertifitseeritud praktikud on otsingutulemites esimesena loetletud, arvestades nende ulatuslikku funktsionaalse meditsiini haridust



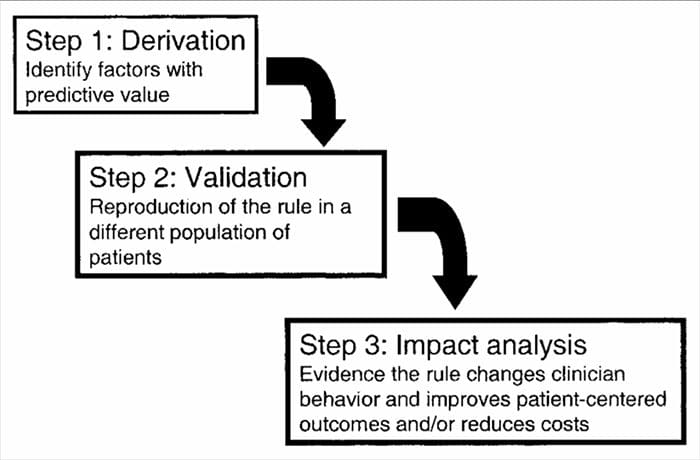




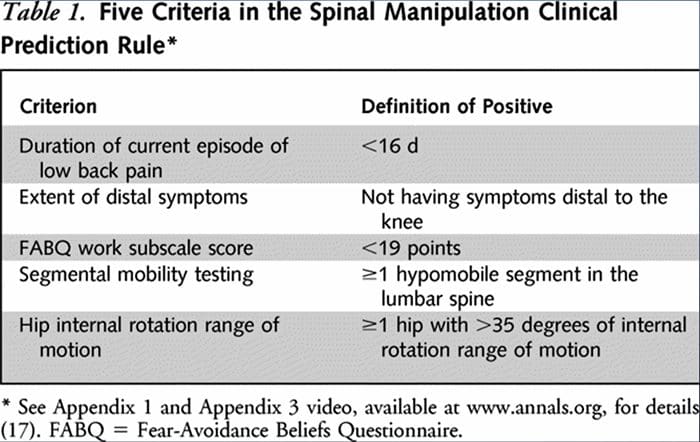
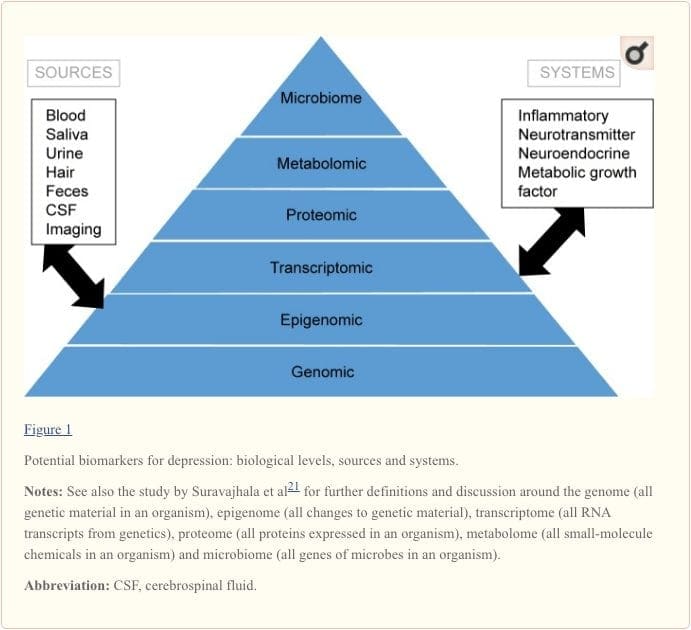





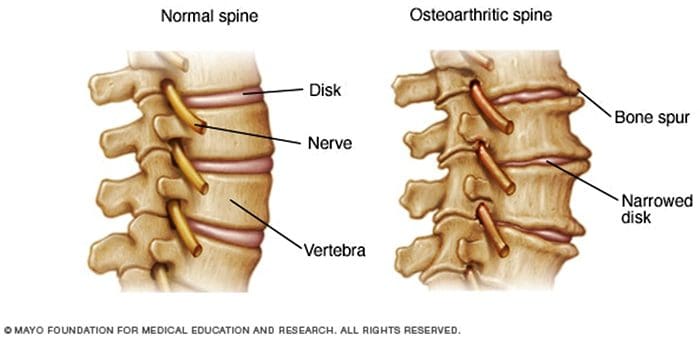
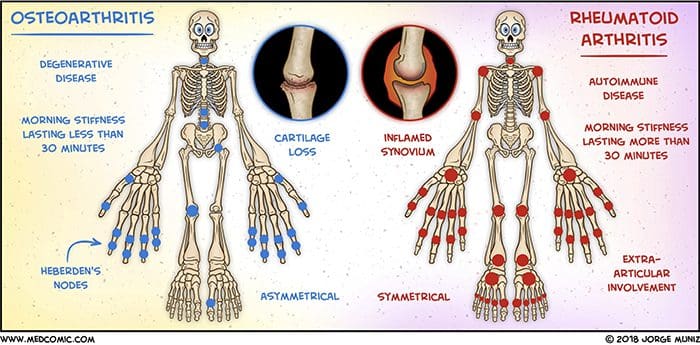 Abstraktne
Abstraktne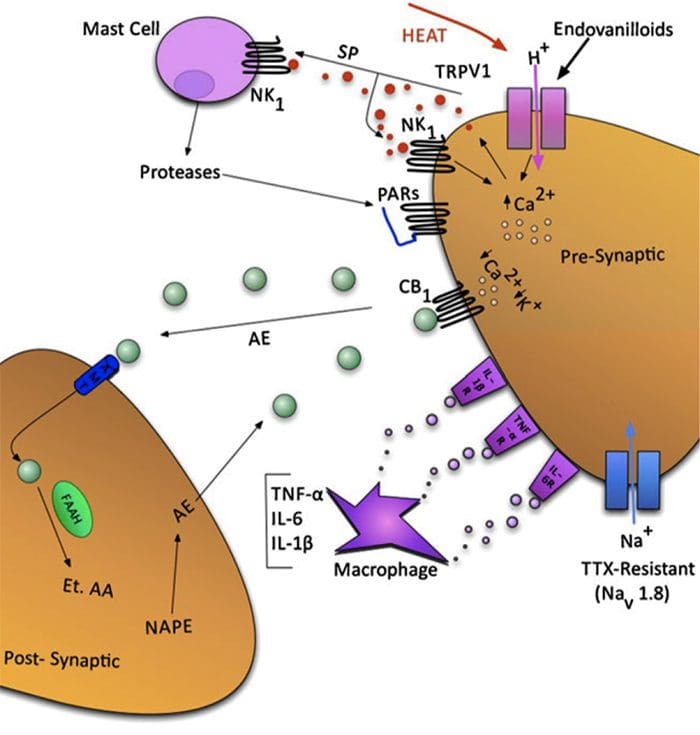
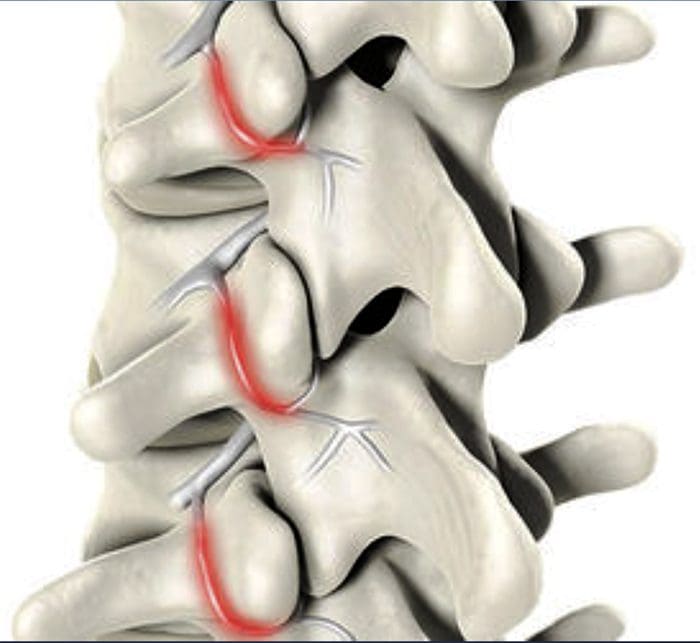
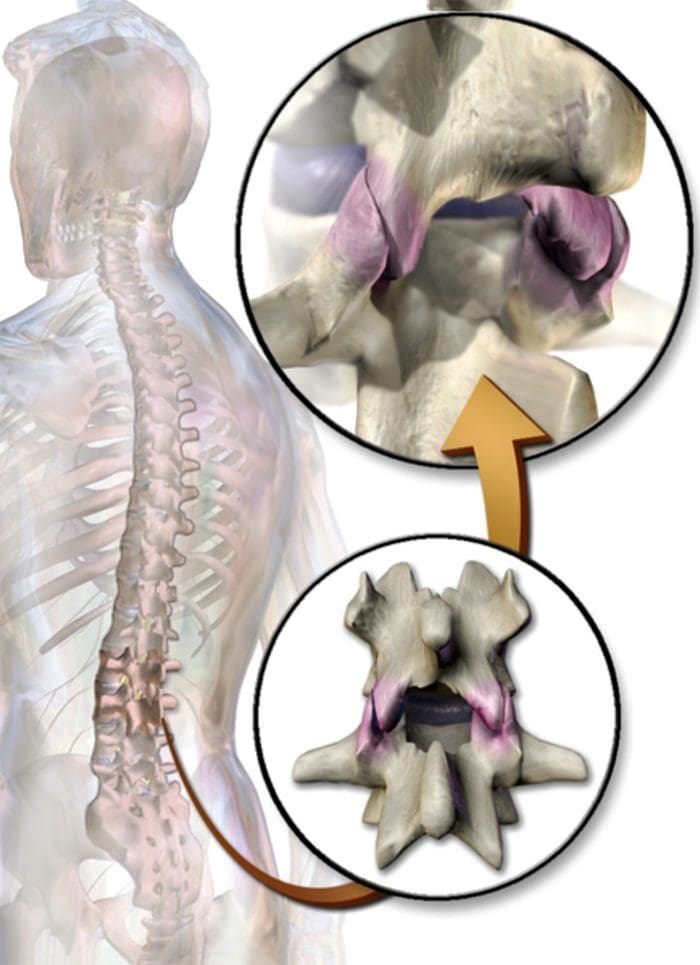
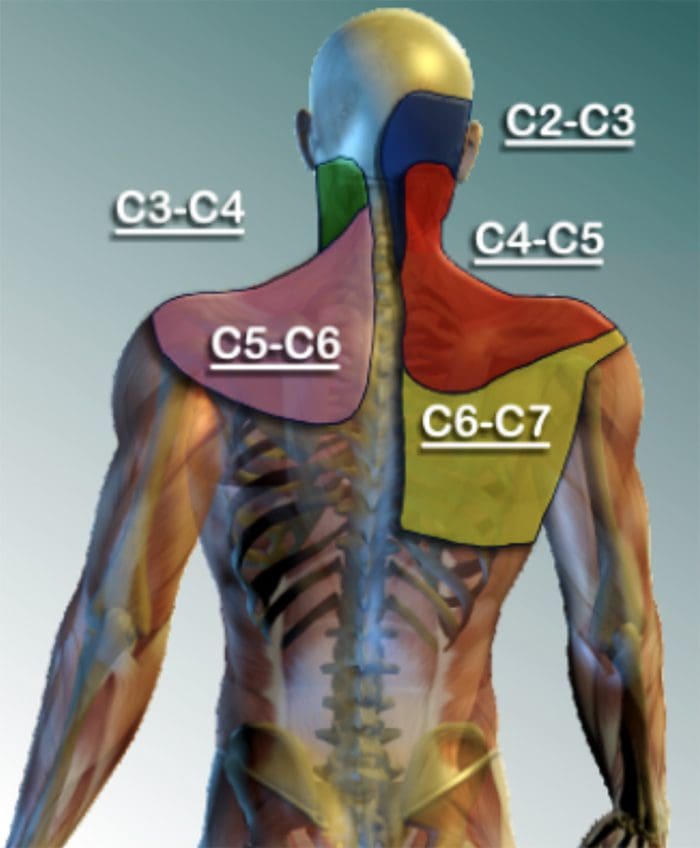
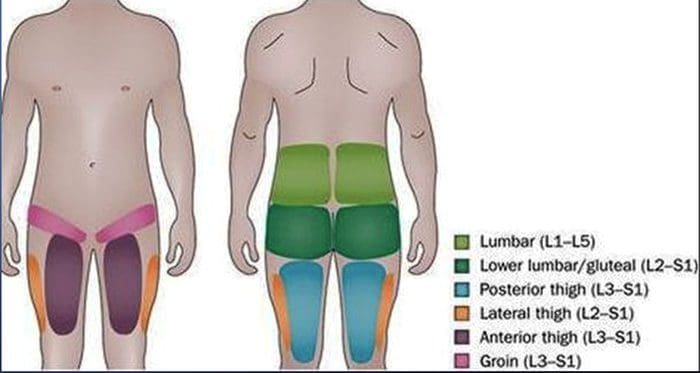

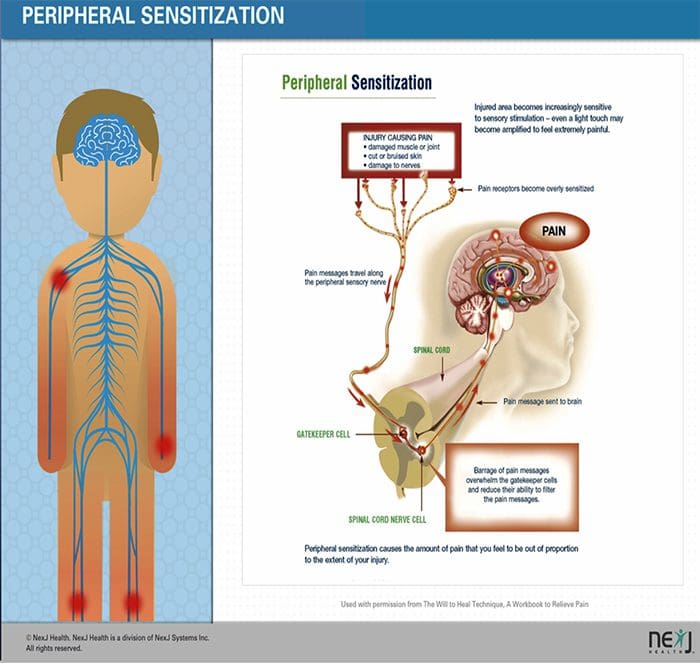
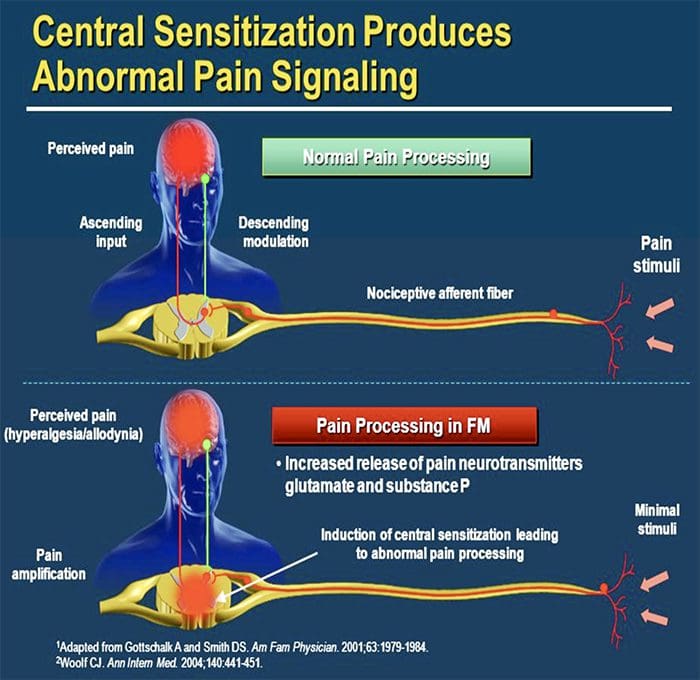


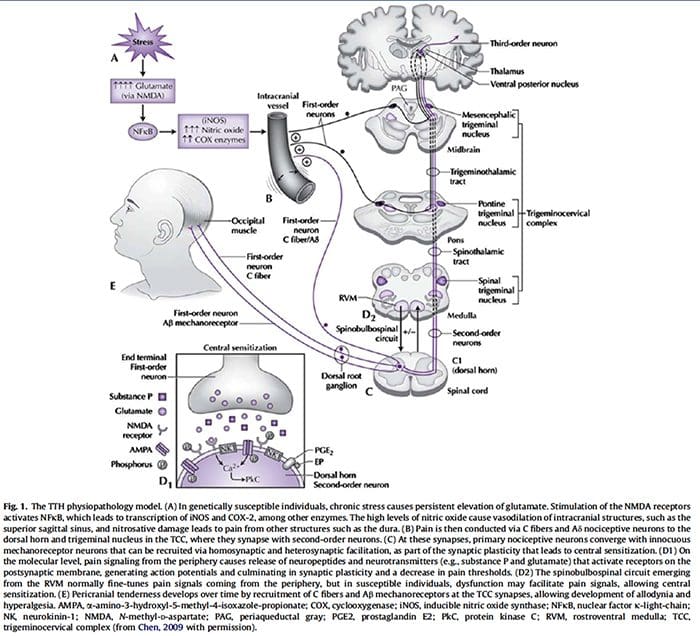

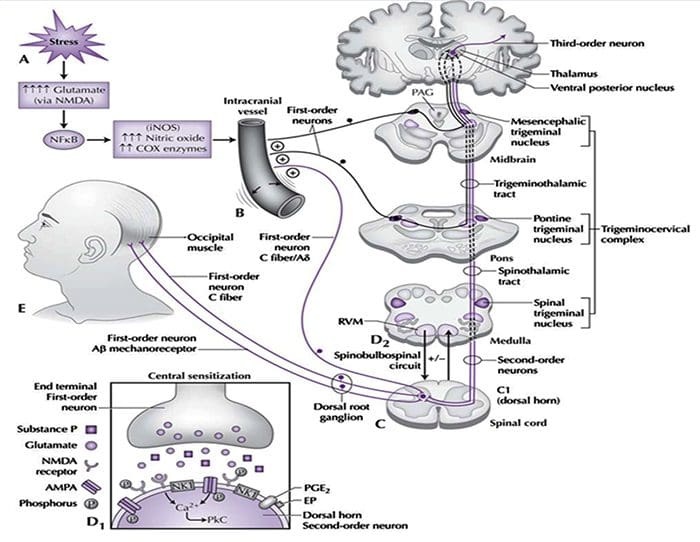




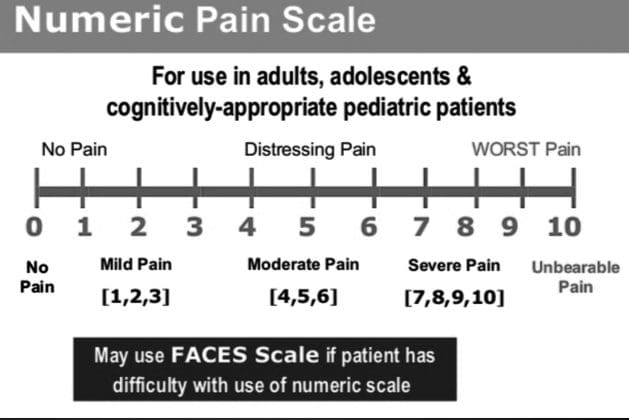
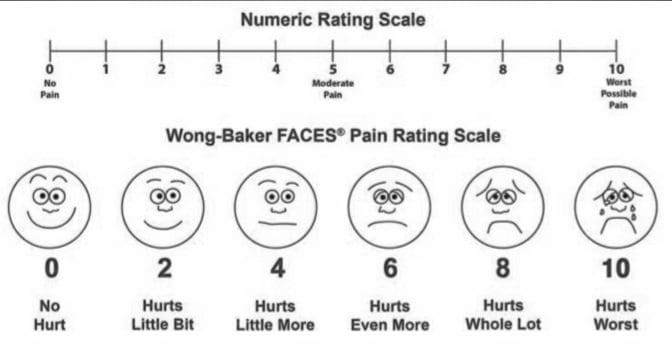








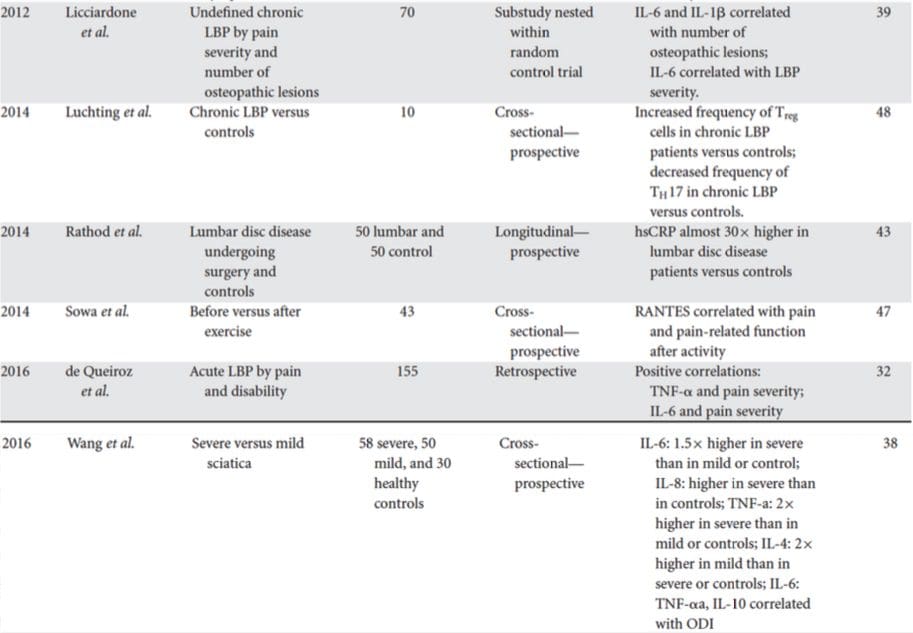


 viited:
viited:
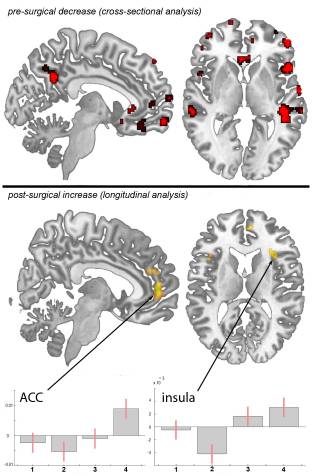
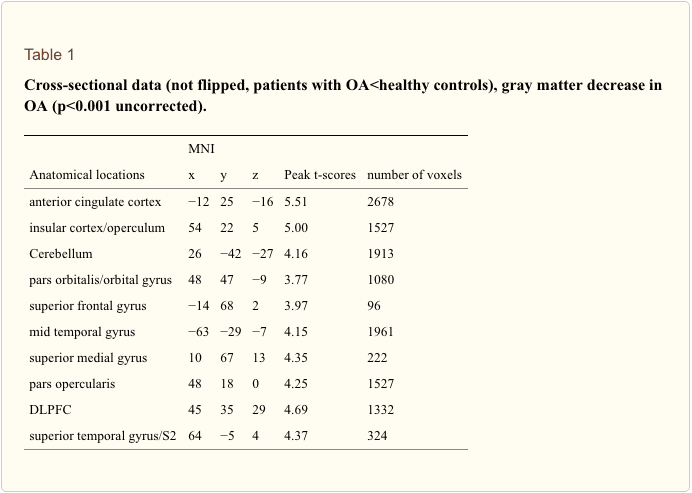
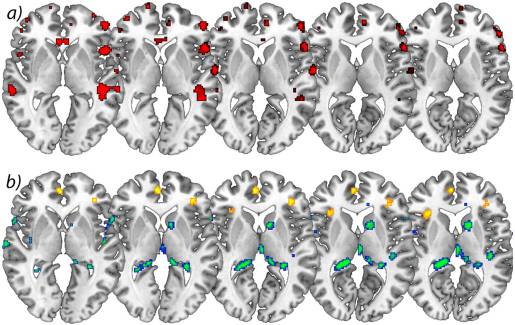
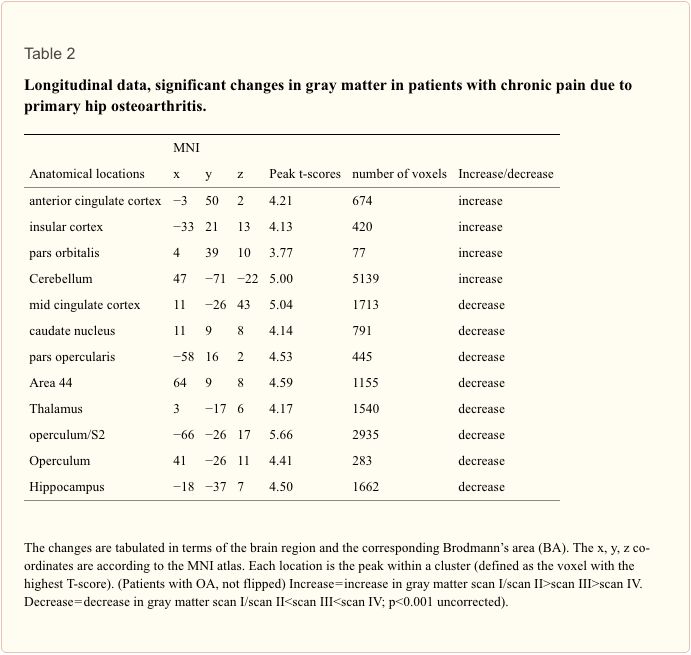
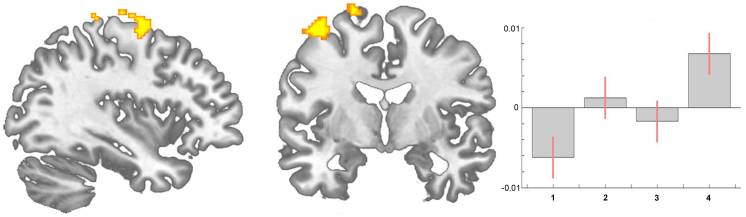
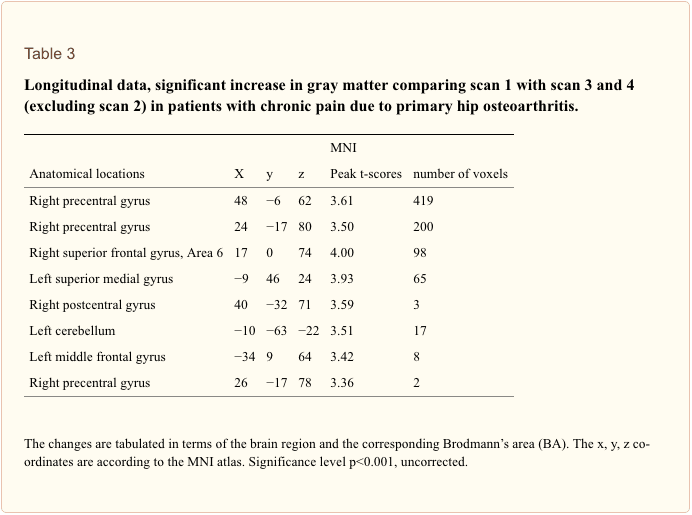

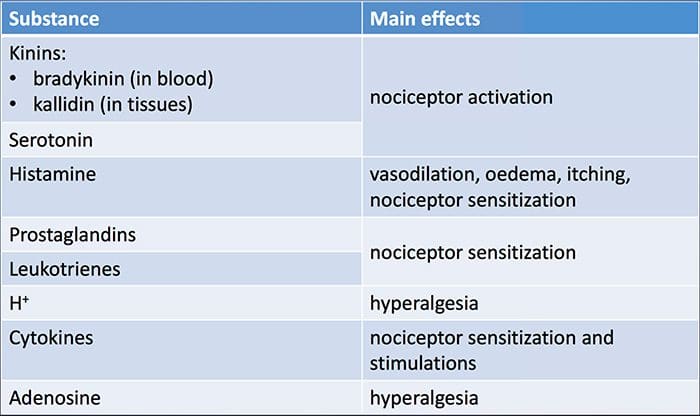
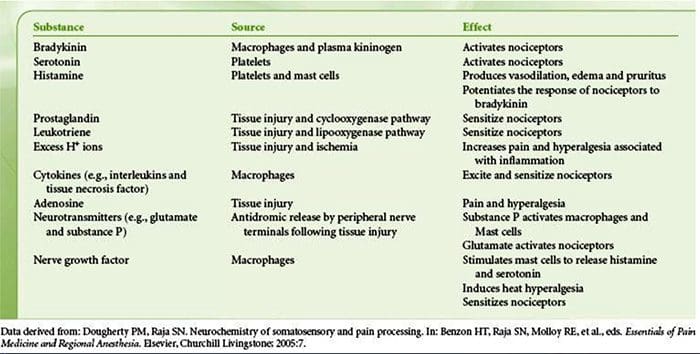
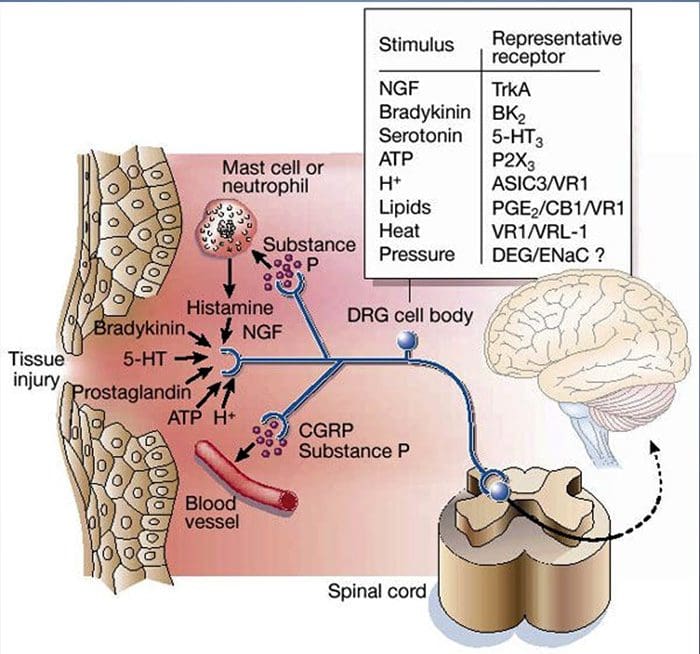
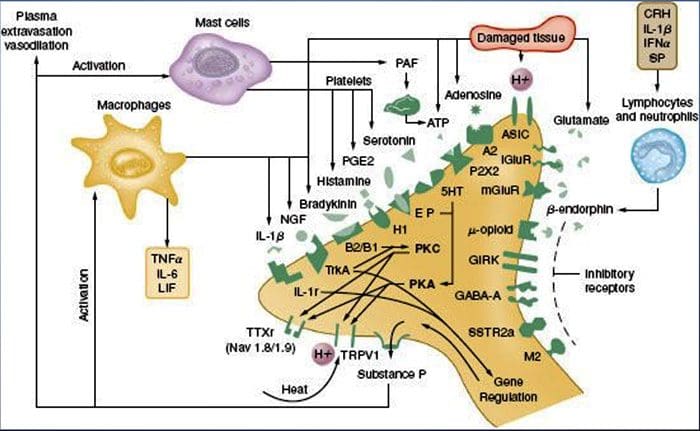 Miks mu õlg valutab? Ülevaade õlavalu neuroanatoomilisest ja biokeemilisest alusest
Miks mu õlg valutab? Ülevaade õlavalu neuroanatoomilisest ja biokeemilisest alusest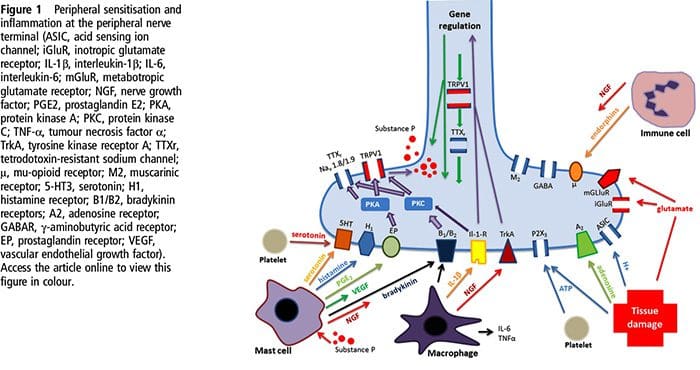 NGF-l ja transientse retseptori potentsiaalse katioonkanali V-alamperekonna V liikme 1 (TRPV1) retseptoril on sümbiootiline seos põletiku ja notsitseptori sensibiliseerimise osas. Põletikulises koes toodetud tsütokiinid suurendavad NGF tootmist. 19 NGF stimuleerib nuumrakkude poolt histamiini ja serotoniini (5-HT3) vabanemist ning sensibiliseerib ka notsitseptoreid, muutes võib-olla A? kiududest nii, et suurem osa neist saab notsitseptiivseks. TRPV1 retseptor esineb primaarsete aferentsete kiudude alampopulatsioonis ja seda aktiveerivad kapsaitsiin, kuumus ja prootonid. TRPV1 retseptor sünteesitakse aferentse kiu rakukehas ja see transporditakse nii perifeersesse kui ka tsentraalsesse terminali, kus see aitab kaasa notsitseptiivsete aferentide tundlikkusele. Põletiku tagajärjel tekib perifeerselt NGF, mis seondub siis notsitseptori klemmides oleva türosiini kinaasi retseptori 1. tüüpi retseptoriga, seejärel transporditakse NGF rakukehasse, kus see viib TRPV1 transkriptsiooni ülesreguleerimiseni ja sellest tulenevalt suurenenud notsitseptori tundlikkus. 19 20 NGF ja ka teised põletikulised vahendajad sensibiliseerivad TRPV1 sekundaarsete messenger-radade mitmekesise hulga kaudu. Arvatakse, et perifeersete notsitseptorite tundlikkuses osalevad ka paljud teised retseptorid, sealhulgas kolinergilised retseptorid, a-aminovõihappe (GABA) retseptorid ja somatostatiini retseptorid.
NGF-l ja transientse retseptori potentsiaalse katioonkanali V-alamperekonna V liikme 1 (TRPV1) retseptoril on sümbiootiline seos põletiku ja notsitseptori sensibiliseerimise osas. Põletikulises koes toodetud tsütokiinid suurendavad NGF tootmist. 19 NGF stimuleerib nuumrakkude poolt histamiini ja serotoniini (5-HT3) vabanemist ning sensibiliseerib ka notsitseptoreid, muutes võib-olla A? kiududest nii, et suurem osa neist saab notsitseptiivseks. TRPV1 retseptor esineb primaarsete aferentsete kiudude alampopulatsioonis ja seda aktiveerivad kapsaitsiin, kuumus ja prootonid. TRPV1 retseptor sünteesitakse aferentse kiu rakukehas ja see transporditakse nii perifeersesse kui ka tsentraalsesse terminali, kus see aitab kaasa notsitseptiivsete aferentide tundlikkusele. Põletiku tagajärjel tekib perifeerselt NGF, mis seondub siis notsitseptori klemmides oleva türosiini kinaasi retseptori 1. tüüpi retseptoriga, seejärel transporditakse NGF rakukehasse, kus see viib TRPV1 transkriptsiooni ülesreguleerimiseni ja sellest tulenevalt suurenenud notsitseptori tundlikkus. 19 20 NGF ja ka teised põletikulised vahendajad sensibiliseerivad TRPV1 sekundaarsete messenger-radade mitmekesise hulga kaudu. Arvatakse, et perifeersete notsitseptorite tundlikkuses osalevad ka paljud teised retseptorid, sealhulgas kolinergilised retseptorid, a-aminovõihappe (GABA) retseptorid ja somatostatiini retseptorid.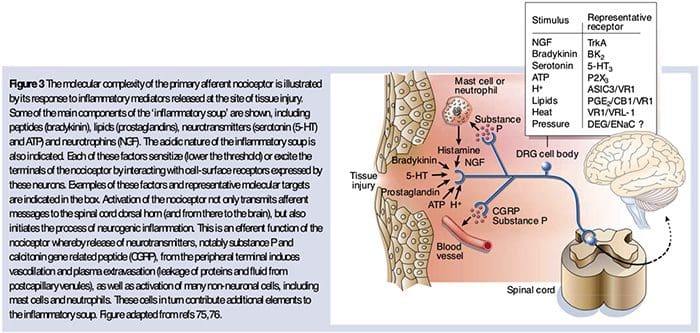 Nociceptoride neurokeemia
Nociceptoride neurokeemia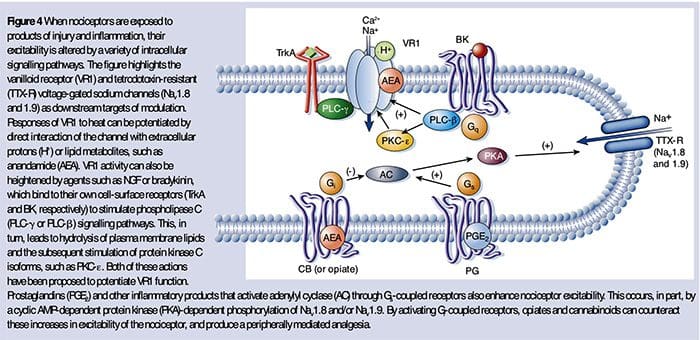 Rakulised ja molekulaarsed valumehhanismid
Rakulised ja molekulaarsed valumehhanismid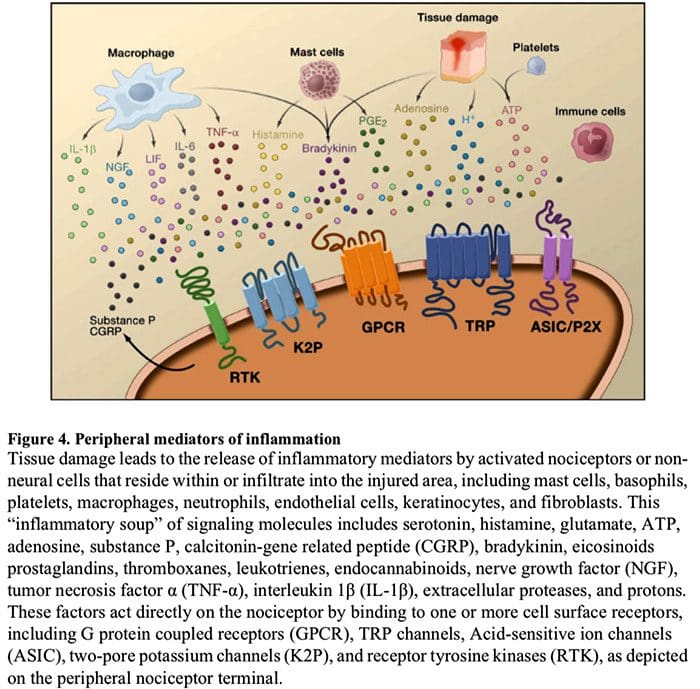
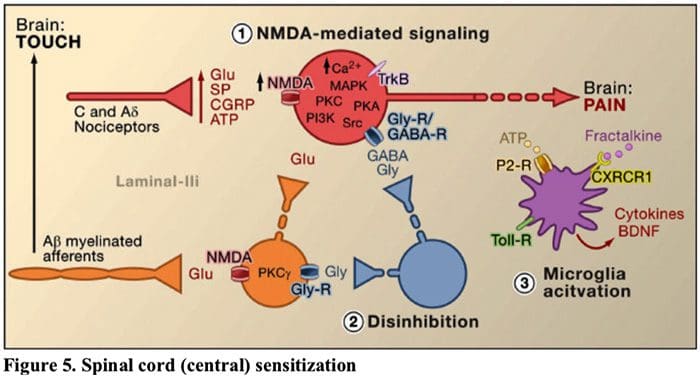 Joonis 5. Seljaaju (keskne) sensibiliseerimine
Joonis 5. Seljaaju (keskne) sensibiliseerimine